年度別活動報告
年度別活動報告書:2004年度
脊椎動物の頭部神経系はどのように部域化されるのか
橋本主税(主任研究員)
辻咲織(大学院生)
山口真未(大学院生)
永友寛一郎(大学院生)
外谷雅範(大学院生)
村戸康人(大学院生)
皐裕美(研究補助員)
はじめに
脊椎動物の頭部神経系の構造は種を越えて高度に保存されており、またその構造を作り出す仕組みも共通の分子機構によって担われているが1)、実際の頭部神経系を見ると、その大きさや形などが大きく異なった結果、異なる種として存在するわけで、このような種間における類似性と相違性を生み出す分子の仕組みを知ることも、構造と機能の進化を知るアプローチとして重要である。
脊椎動物では、オーガナイザーと呼ばれる内中胚葉性の機能領域の働きにより、未分化外胚葉組織が、部域化された神経組織へと誘導される2)。発生が進むとオーガナイザーはさらに詳細に領域化され、それに伴って頭部神経組織の領域化もすすむ。オーガナイザーや神経組織が領域化される過程は、母性由来の緩やかな位置情報からシャープな位置情報の獲得であると言える。これは生きものの形づくりの過程で頻繁に利用されている重要な方法論であるため、普遍に存在する秩序の形成を考える上で非常に興味深い3)。初期発生過程において何らかのパターンを作る場合、一般に、パターンが存在しない場所に、特定の起点からの「物差し」を作る必要がある。これはどのようにして成立しているのだろうか?たとえば特殊な波動方程式4)などによる説明が知られているが、その分子的実体は明らかになっていない。また、大まかに規定された物差しの情報をさらにきめ細かな物差しの情報へと変換する機構に関しての分子レベルでの解析は明確にされていない。
我々は、アフリカツメガエル(以下ツメガエル)を実験材料として形態学と分子生物学的手法の両面から研究を遂行している。ツメガエルの頭部神経形成過程では、部域化されているオーガナイザー組織が予定頭部神経組織に接し、自己の持つ位置情報を外胚葉組織に与えることで領域化された神経を誘導する5)。この「頭部神経の領域化」を制御する機構は、広範囲の組織(細胞)運動を伴いながら組織の性質が変わり(分化)、周りの組織との間に明瞭な境界を規定する一方で独自の形態を作り上げる(形態形成)という、生物の形づくりの時に一般に見られる現象を全て含んでいると考えられる。したがってツメガエルの脳がどのような仕組みで形づくられていくのかについての全体像を明らかにすることで、生きものの高次構造形成に潜在する普遍的な現象に到達できるのではないかと期待するのである。
結果と考察
1)Xhairy2b遺伝子は、複数の機能を持つ単一分子として発生過程の胚に大まかなパターンを作らせる
胚の背側帯域に形成される初期のオーガナイザーは、まず腹側組織との差別化がなされており、また、オーガナイザー内部でも内胚葉領域と中胚葉領域におおまかに部域化が行なわれている。
我々は既に、予定脊索前板に特異的に発現する遺伝子Xhairy2b 6)が体幹部を誘導できるが、頭部を誘導できないことを明らかにしている。この事実は、現在までに提示されている「脊索前板は頭部誘導能を持つ」という説とは異なり 7)、「(予定)脊索前板は体幹部の誘導に働く」ことを示す。またXhairy2b遺伝子は、複数のオーガナイザー特異的な遺伝子群の発現を互いに誘導できること、脊索前板に隣接する他のオーガナイザー領域である内胚葉や脊索に特異的な遺伝子の発現を抑制することも明らかにした 8)。これらの事実を統合すると、オーガナイザー内部の特定の細胞に発現したXhairy2bは、その細胞では脊索前板特異的遺伝子の発現を保証するとともに、隣接する組織への分化転換を積極的に抑制することにより、予定脊索前板のアイデンティティーの確立に重要な働きをしていると考えられる。(図1) しかしこれらの事実からは、オーガナイザー(背側)領域が腹側と差別化される機構やオーガナイザー内部領域が内胚葉組織と中胚葉組織に差別化される機構などの「大まかな部域化」と、脊索前板のさらに前方部領域の「厳密な部域化」という二つの重要な機構は理解できない。
図1:初期原腸胚におけるXhairy2b遺伝子の役割
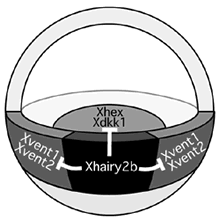
初期原腸胚の背側中胚葉(オーガナイザー)に発現するXhairy2bは、腹側中胚葉(Xvent1,2)や前方内胚葉(Xhex, Xdkk1)など隣接する領域の形成を積極的に抑制することで、背側中胚葉としてアイデンティティーを確立している。
我々は、Xhairy2b遺伝子を発現させた場所でfollistatinなど背側特異的遺伝子の発現が活性化され、Xvent1などの腹側特異的遺伝子の発現が抑制されることを見いだした。この現象を詳細に解析したところ、Xhairy2b遺伝子を強制発現させた場所の周辺領域で背側特異的遺伝子ののひとつであるchordinの発現が誘導されることが分かった。これらの結果から、Xhairy2b遺伝子を腹側組織に強制発現させると、発現領域に重なってfollistatinなどが、Xhairy2b遺伝子の発現場所とは重ならずその周辺領域にchordinが誘導され、そのさらに外側にXvent1などが同心円のように発現するモデルが考えられる(図2)。
図2:Xhairy2b遺伝子は均一な空間に「物差し」を形成する。

Xhairy2b遺伝子を腹側領域に発現させることで、背側前方に特異的な遺伝子(follistatin)・背側後方に特異的な遺伝子(chordin)・腹側に特異的な遺伝子(vent)の順に同心円状の発現領域を形成することができる。
一般に、脊椎動物の背腹軸決定過程においては腹側性質が優位であり、背側化活性(すなわちオーガナイザー)が作られない場合には胚は全体が腹側となることが知られている。また、オーガナイザー自体が背腹軸そして前後軸に沿ったパターン形成を受け、そのパターンが胚全体のパターンを決定づけることも知られている。今回Xhairy2b遺伝子の強制発現によって見られたおおまかなパターン形成は、原則的に腹側として形成されたツメガエル胚の一点にXhairy2b遺伝子が発現することで、前方部背側領域・後方部背側領域・腹側領域の順に胚に大まかな物差しをおいていると考えられる。さらにXhairy2b遺伝子は、胚の再前方組織の形成を抑えることも知られており、この遺伝子が胚の背腹軸と前後軸のおおまかなパターンを決めるのに重要であることが推測できる。
図3:Xhairy2bが属するHESファミリー蛋白質のドメイン構造。

また、Xhairy2b遺伝子によるパターン形成制御を解析している途中で興味深い現象を見いだした。bHLH型の抑制性転写因子であるXhairy2b分子内には、DNA結合ドメイン・二量体形成ドメイン・機能未知ドメイン・転写抑制に関与するドメインなどが進化的に保存されている(図3)。一般にこれらのドメインのはたらきは、転写因子としてのXhairy2bのはたらきに必須である。ところが、Xhairy2b蛋白質のC末端ドメイン単独で腹側特異的遺伝子の発現抑制と、一部の背側特異的遺伝子の発現誘導を行なうことが分かったのである。この蛋白質はDNA結合能を有さず、したがってもはや転写因子としてのはたらきを持たないため、何らかの構造蛋白質として働くことが推測できる。詳細な解析から、このC末端ドメインによって発現が誘導される遺伝子と抑制される遺伝子、そして転写因子としてのXhairy2b蛋白質が発現誘導する遺伝子と抑制する遺伝子はそれぞれ異なり、これらが互いに密接に絡み合いながら、結果として、図2で示した同心円状のパターンを描き出すと結論付けられた。Xhairy2b蛋白質の各ドメインがそれぞれ具体的にどのようなはたらきをしているのかについて現在解析中である。
2) Xhairy2b遺伝子は脊索前板の前方領域を厳密に規定する
Xhairy2bによっておおまかに領域化されたオーガナイザーを詳細に見ると、領域の境界部分では隣り合う領域の細胞が入り交じって存在し、そこには明瞭な境界線が見いだせない。しかし、オーガナイザーは、最終的に咽頭内胚葉・内中胚葉・脊索前板・脊索などに分かれ、さらに脊索前板は前方部と後方部など異なる組織に分かれ、それぞれの組織間の境界ははっきりしてくる事が見て取れる。この詳細な領域化がどのように成立するかについては現時点では明らかではない。
図4:アンチセンスオリゴによるXhairy2bの機能阻害。
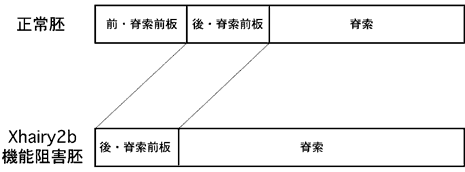
Xhairy2bの機能阻害により前方脊索前板の形成が阻害され、脊索前板の前後境界や脊索前板と脊索との境界が前方部に移動する。
オーガナイザーにおけるXhairy2b遺伝子の機能をアンチセンスオリゴを用いて人為的に阻害したところ、脊索前板前方部の形成が特異的に抑制され、脊索前板の前方と後方の境界が前方にシフトしていた(図4)。また、Xhairy2b遺伝子は前方部脊索前板特異的な遺伝子群の発現を互いに誘導でき、さらには後方部脊索前板に特異的な遺伝子の発現を抑えることも明らかとなった。これらの結果から、Xhairy2b遺伝子は、オーガナイザーに由来する組織のうちの脊索前板前方部領域を厳密に規定することが示された(図5)。Xhairy2bを含むHES遺伝子ファミリーは種々の発生現象において境界形成を厳密に規定することが知られているので、この脊索前板の前後境界形成機構をより詳細に解析することにより、境界形成機構を一般に論じることができるかも知れない。
図5:初期神経胚におけるXhairy2b遺伝子の役割
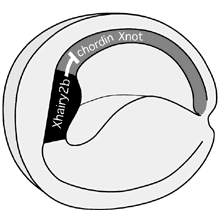
初期神経胚でXhairy2b遺伝子は、後方部脊索前板(chordin)と脊索(chordin, Xnot)の形成を積極的に阻害すつことにより、前方部脊索前板のアイデンティティーを確立している
さらにここで興味深いことは、Xhairy2bの機能阻害によって脊索前板前方領域の形成を阻害した胚では、前脳の特定部域(視床下部)の形成が阻害されていた。オーガナイザー由来の組織が脳のパターンを制御することは言われているが、このように脊索前板前方領域という特定の部分が予定視床下部の形成に必須であるという詳細な解析は他になく、これをさらに解析することで、脳のパターン形成の仕組みの一端を明らかにできることが期待される。
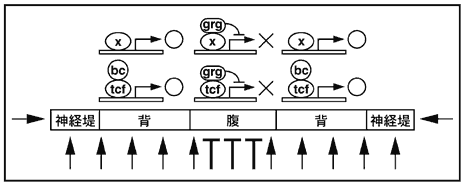
3) Wnt/βカテニンシグナルが間脳の背腹軸を決定する
オーガナイザーはそれ自身が領域化され、各オーガナイザー領域からの位置情報が頭部神経組織の位置情報となる。我々は、予定間脳領域の背腹決定に注目して、その領域化を制御するシグナルについて解析を進めてきた。具体的には、Wntシグナル経路では、最終的な転写因子であるLEF/Tcfに普段はGrouchoが結合して転写を積極的に抑制しているが、一度Wntシグナルが活性化されるとβカテニンがGrouchoに代わりLEF/Tcfと結合し、その標的遺伝子の転写を促進すると考えられている。我々はAxin/Axilによるシグナル経路の不活性化や、βカテニンによる活性化を行ない、間脳の背腹軸決定に影響があることを確認した。次に、βカテニンあるいはGrouchoとの結合能をそれぞれ欠く変異LEF/Tcfの効果を解析したところ、βカテニンとの結合能を欠くLEF/Tcfを強制的に発現させると背側領域での活性が予想されるWntシグナルに応答できないため、結果として間脳全体の腹側化が起こり、逆にGrouchoとの結合能を欠くLEF/Tcfの存在によって間脳は全体として背側化すると事が明らかとなった。これらの結果に基づいて、原腸胚期の予定間脳領域ではWntシグナルが活性化されることにより背側の性質を獲得し、Wntシグナルが抑制されることによって腹側の性質を獲得するという図6に示すモデルを提唱した9)。このモデルでWntシグナルは周囲の外胚葉や中胚葉に由来するとしているが、厳密には今後解明されるべき問題である。ここで特筆すべき事は、Wntシグナルはこれまで神経領域の後方化シグナルとして知られていたことである。それが前方神経組織の胚腹軸を決める仕組みにも使われてきたことは非常に興味深く、さらに前後軸形成と胚腹軸形成が時間的空間的にどのように制御されてきたかについて新たな疑問を提唱する。また、前章でも言及したが、前方部脊索前板が視床下部の形成を制御する。視床下部は間脳腹側領域に由来する組織であり、このWntシグナルの関係性も興味深い。
図6:原腸胚期の予定間脳領域における背腹決定モデル
間脳で、Wntシグナルが存在する場所(矢印)に背側領域が、Wntシグナルの存在がない場所(丁字)に腹側領域が形成される。
本研究により、間脳の背腹を決めるシグナルの一端が垣間見えてきたが、あくまでも「間脳の背腹」の決定機構に過ぎない。なぜ間脳以外の領域の背腹はWntによって影響を受けないのか?あるいはそもそも間脳を決める機構は何か?について解答がもたらされなければ脳の領域化を理解することはできない。まだまだスタートラインに立ったところではあるが、いままで奥の見えなかった暗闇に一筋の光が差し込んだことは事実であろう。
おわりに
あらかじめパターンの存在しない空間に、大まかな物差しを決め、さらにそこから詳細に領域化される機構について、特にオーガナイザーの形成と部域化並びに初期神経のパターン形成においては、Xhairy2bの働きによってひとつのモデルを提唱できたと考えている。しかし、発生現象を分子生物学的に考えた時にはパターンを形成するものは細胞であり、それぞれの細胞運命を決める分子機構が理解されなくてはならない。いわば、人文字を形成する時のように、個人の意志を越えて全体を動かす総合的な仕組みが何かを求めなければ、生きものの形が出来上がる仕組みを知ることはできない。機械論的ではなく、確率論的にさえ見える個々の細胞の振る舞いを分子によって解き明かすことはこれからの発生学において重要な課題のひとつであり10)、本研究の最終目的のひとつにこの課題への解答があることは間違いのないことであるが、現時点ではそのヒントすら得られていない。これは「いい加減さ」や「大ざっぱさ」を許容できる「しなやかさ」を意味し、決して機械には真似のできない生きものが生きものとして存在する本質的現象である。
最後に、新しく得られている結果から将来的に魅力のある事に触れる。ツメガエルは偽4倍体であり、同一遺伝子を複数種類持つことが知られている。Xhairy2b遺伝子も同様に、Xhairy2aと言う遺伝子が知られている。Xhairy2aとXhairy2bはアミノ酸配列にして92%以上相同であり、転写因子としての機能も同等であるとされているが、特異的に機能阻害を行なった場合、その現象に応じてXhairy2bのみが機能している事が見いだされている。現在までに両者の発現パターンは等しいことが知られているので、この相違は遺伝子そのものの違いに由来するはずである。ではその違いとは何であろうか?現時点では何も分からないが、生体内においてXhairy2b遺伝子は期待されるよりも長い蛋白質を作っているらしく、その蛋白質は蛋白質をコードする領域の上流に位置する短いORFとの融合蛋白質であるらしい。このような、翻訳の段階での制御によって発生現象に詳細な調節を行なう例はそれほど知られていないが、ここで発生現象を制御する新しい機構を見ている可能性も考えられる。今後、この現象にも着目して解析を進めたい。
参考文献
1) Holley, S.A. (1997) Fish are like flies are like frogs: conservation of dorsal-ventral patterning mechanisms. BioEssays 19: 281-284
2) Spemann, H. and Mangold, H. (1924) Wilhelm Roux's Arch. 100: 599-638
3) Holloway, D.M., Reinitz, J., Spirov, A. and Vanario-Alonso, C.E. (2002) Sharp borders from fuzzy gradients. Trends Genet. 18: 385-387
4) Shoji H, Iwasa Y, Kondo S. (2003) Stripes, spots, or reversed spots in two-dimensional Turing systems. J Theor Biol. 224(3):339-50.
5) Koide, T., Umesono, K and Hashimoto, C (2002) When does the endomesoderm meet the anterior-most neuroectoderm during Xenopus gastrulation? Int. J. Dev. Biol. 46: 777-783
6) Tsuji, S., Cho, K.W.Y. and Hashimoto, C. (2003) Expression pattern of a basic helix-loop-helix transcription factor Xhairy2b during Xenopus laevis development. Dev Genes Evol. 213: 407-411
7) Shibata M., Itoh M., Ohmori S.Y., Shinga J. and Taira M. (2001) Systematic screening and expression analysis of the head organizer genes in Xenopus embryos. Dev Biol. 239:241-56.
8) Yamaguti, M., Cho. K. W. Y. and Hashimoto, C. (2005) Xenopus hairy2b specifies anterior prechordal mesoderm identity within Spemannユs organizer. Submitted to Mech. Dev.
9) Tsuji, S. and Hashimoto, C. (2005) Choice of either b-catenin or Groucho/TLE as a co-factor for Xtcf-3 determines dorsal-ventral cell fate of diencephalon during Xenopus development.
Dev Genes Evol. In press
10) Ko, M.S.H., Nakauchi, H. and Takahashi, N. (1990) The dose dependence of glucocorticoid-inducible gene expression results from changes in the number of transcriptionally active
templ ates. EMBO J. 9: 2835-2842