年度別活動報告
年度別活動報告書:2008年度
脊索動物と節足動物の共通祖先を理解する 2−1 オオヒメグモの胚発生を支配する分子メカニズムの解明
小田広樹(主任研究員) 秋山-小田康子(奨励研究員)
野田彰子(研究補助員)
春田知洋、金山真紀(大阪大学大学院生)
はじめに
系統的に遠く離れていると考えられている2つの動物グループ、脊椎動物と昆虫において共通に存在する遺伝子が類似した発生現象に関わっている例が多数見つかっている1)。代表的な例としては、体の前後や背腹のパターンを決める遺伝子や眼の分化を制御する遺伝子である。このような発生システムの類似性を根拠として、今から6億年以上前に現在の脊椎動物と節足動物に共通に見られる特徴をもつ動物が誕生しており、その子孫として現在の多様な動物群が現れたと考えられるようになっている。もしこの考えが正しいとすると、どのような変化がそれぞれの動物グループを誕生させたのだろうか? そのような変化を説明できなければ動物の多様性の成り立ちを理解したとは言えない。とりわけ、無脊椎動物から脊椎動物が誕生した過程において起こった変化を調べることは私たちヒトの歴史をたどる意味も持つ。脊椎動物の発生において最も重要な役割を果たす脊索の起源は何か? 脊椎動物の体節は無脊椎動物に見られる繰り返し構造と同じ起源を持つのか、それとも脊椎動物の系統で独自に進化したのか? これらの問題を解くためには、脊椎動物が誕生する以前の無脊椎動物の状態を如何に知るかが重要である。そのために私たちは、節足動物門の鋏角類に属するオオヒメグモ (Achaearanea tepidariorum) を新しいモデル動物として開発してきた。
私たちがオオヒメグモに注目する理由は主に以下の5つである。1)系統的に見て昆虫と脊椎動物の間に位置すること。2)脊椎動物と同じように体軸を調節的に発生させ、細胞間の相互作用に大きく依存した発生システムをとっていること2),3)。3)実験を遂行する上で利便性があり、今後の技術開拓の可能性も十分にあること。4)微細形態が脊椎動物のものに酷似したタイトジャンクションを持っていること4)。5)ショウジョウバエのゲノムにはないが、脊椎動物とオオヒメグモのゲノムに共通して存在する遺伝子が多数存在すること。
これまでの研究で私たちは、ディピーピー-ソグ、デルタ¬-ノッチ、ヘッジホッグ¬-パッチトのシグナル経路がオオヒメグモ胚の体軸形成に重要な役割を果たしていることを明らかにした5),6)(論文投稿中)。ソグの機能解析を通じて、クモの腹側正中線外胚葉の特異化の仕組みが脊椎動物の背側の正中線組織(脊索/神経底)の特異化の仕組みに類似していることも示してきた。昨年度の解析では、ヘッジホッグシグナル経路の正常な働きが前後軸・背腹軸形成、尾部形成、体節形成のすべてに必須であることが分かってきており、本年度はこのシグナル経路を主軸に置いて研究を推し進めた。ヘッジホッグシグナルと体軸形成との関係が示されている動物種は、現在までのところオオヒメグモだけである。このオオヒメグモの状態が節足動物にとって特殊な状態なのか、それとも祖先的な状態なのかは他の動物種での研究を待たなければならないが、脊椎動物や昆虫の付属肢における軸の発生で共通にヘッジホッグシグナルが主要な役割を果たしていること7)を考えると、軸やパターンを生み出す普遍的な(祖先的な)仕組みをオオヒメグモ胚を使って解明できるかもしれない。ヘッジホッグシグナル経路の働き方は非常にダイナミックであり、構成因子の作用の仕方も複雑である8),9)。しかも、他のシグナル経路とも巧みに連係しているために解析が難しい。シンプルな細胞構成でかつ比較的広い“場”をもち、複数のシグナル経路が同時に活性化しているオオヒメグモ初期胚は、ヘッジホッグシグナル経路の研究の新しいモデルシステムになりうるのではないかと期待される。
結果と考察
(1)尾葉形成に関わる遺伝子の探索
クモの胚帯の後端部は伸長しながら体節が形成される。この後端領域は尾葉と呼ばれ、もともと初期胚の原口を取り囲む領域から形成される。これまでの解析から、胚盤期に原口付近から胚盤の周縁に向かって対称に広がる発現がデルタなどの複数の遺伝子について観察されており、この遺伝子発現が尾葉形成と深く関わっていることが分かっている。さらに、パッチトRNAi胚ではその原口周りからの遺伝子発現が全く起こらず、尾葉も全く形成されないことが分かっており、このことを利用して尾葉形成に関わる遺伝子を網羅的に探索できるのではないかと考えた。
これまでのEST解析により、14,444本の重複しないDNA塩基配列が得られているが、この配列情報を用いて24,000種類のオリゴDNAプローブ(40mer)をデザインし、in situ合成方式のマイクロアレイを作製した。そして、[胚盤期後期の正常胚]と[同じステージのパッチト、ディピーピーの二重RNAi胚]からトータルRNAを精製し、マイクロアレイ解析によって両者の間の遺伝子発現の比較を行った(図1)。[パッチトRNAi胚]ではなく、[パッチト、ディピーピーの二重RNAi胚]を解析に用いた理由は、ディピーピーシグナルに支配される遺伝子も同時に検出したいと考えたからである。
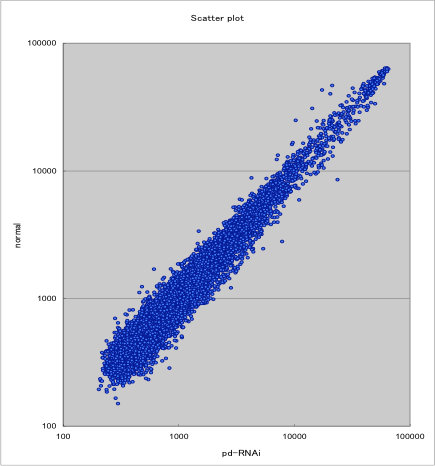 |
図1 マイクロアレイ解析の結果
各スポットについて[正常胚(normal)]と[パッチト、ディピーピーの二重RNAi胚(pd-RNAi)]のシグナルの蛍光強度をプロットした。
シグナルの蛍光強度の比率[パッチト、ディピーピーの二重RNAi胚]/[正常胚]が0.6以下のスポットを“ポジティブ”と判定し、134クローンの候補を得た。これらの中には、ディピーピーやウィント5など実験前から期待された遺伝子も確かに含まれており、マイクロアレイ解析の妥当性が示された。134個の候補クローンすべてについて、in situハイブリダイゼーションによる染色実験を行い、領域特異的な発現パターンを示す28クローンに絞り込んだ。それらの多くは、胚盤期の原口周辺領域や胚帯期の尾葉で特異的な発現を示した。現在、その28クローンすべてについてRNAiによる遺伝子機能抑制実験を行っている。
マイクロアレイ解析と並行して、ハイスループットin situハイブリダイゼーションによる遺伝子スクリーニングも実施している。ESTクローンを基に作製した各プローブに対して胚盤期と胚帯期の2つの胚を染色している。これまでに1584個のプローブによる染色を完了した。
(2)オオヒメグモの体軸形成におけるヘッジホッグシグナル経路の機能解析
前年度までの解析により、オオヒメグモの体軸形成においてヘッジホッグシグナル経路が極めて重要な役割を果たしていることが分かっていた。ヘッジホッグシグナル経路の仕組みは主にショウジョウバエと脊椎動物で解析されているが、多数の構成因子がダイナミックにかつ複雑に関係し合っており、既存のモデルシステムの解析で十分な時空間的解像度が得られているかは疑問である。さらに、ショウジョウバエと脊椎動物で仕組みに重大な違いがあることも指摘されており10),11)、ショウジョウバエの知見をそのまま脊椎動物に適用できるとも限らない。そのような状況において、既存のモデルシステムに比べて比較的単純な細胞構成と比較的広い“場”をもつオオヒメグモの初期胚は、ヘッジホッグ研究の優れたモデルシステムとなる可能性を秘めている。今年度はその可能性を検討するために、これまでに解析したヘッジホッグとパッチトに加えて、新たに3つのヘッジホッグシグナル経路遺伝子スムースンド、キュービタスインタラプタス、サプレッサー・オブ・フューズドの解析を行った。
RNAiによる遺伝子機能抑制実験によって、すべての遺伝子について激しい体軸形成の異常が観察された。スムースンドRNAi胚はヘッジホッグRNAi胚とほぼ同じ表現型を示した。パッチトRNAi胚で見られたようなクムルスの移動の異常はいずれの遺伝子のRNAi胚でも観察されなかった。3つの遺伝子のRNAi胚の表現型は今後、分子マーカーを用いて詳しく解析する予定である。
これまでの結果から注目すべき点は、サプレッサー・オブ・フューズドの機能抑制によってパターン形成に関わる大きな異常が引き起こされたことである。ショウジョウバエにおいて、サプレッサー・オブ・フューズドの単独の突然変異では表現型が全く表れない12)。それに対し、マウスにおいてはサプレッサー・オブ・フューズドの単独の遺伝子破壊がヘッジホッグシグナルに関わる致命的な異常を引き起こす11)。ヘッジホッグシグナル経路においてサプレッサー・オブ・フューズドが果たす役割の重大性だけを見れば、オオヒメグモの状況はショウジョウバエよりもマウスの状況に近いかもしれない。オオヒメグモを使った研究によって、ショウジョウバエでは進化の過程で変質化してしまって見ることができない、普遍的な(祖先的な)仕組みを解明できるのではないかと期待している。
(3)オオヒメグモの頭胸部における体節形成
オオヒメグモの幼生はそれぞれ複数の体節をもつ頭胸部と後体部からなる。後体部の体節は、胚盤の中心領域から生じた胚帯後端部(尾葉)が後方へ向けて伸長するとともに順次形成される。それに対して、頭胸部の体節は胚盤の周縁領域がダイナミックな再配列運動を伴って前後に伸長し、比較的同調的に体節が形成されているように見える。しかし、今年度ヘッジホッグを分子マーカーとして用いて体節形成の様子を詳細に観察したところ、発生中の頭胸部前方領域(頭胸部の前半)において、ヘッジホッグの一つのストライプが2回の分割を経て3つのストライプに変化することが分かった。つまり、少なくとも頭胸部の一部の体節は同調的ではなく、後体部の体節と同じように順次形成されていた。
さらに、胚盤の周縁領域で発現し、体節形成とともに複数のストライプに発現パターンが変化する遺伝子オッドペアードに注目した。この遺伝子の機能をRNAiにより抑制すると、頭胸部前方領域のヘッジホッグのストライプの分割が起こらなくなった(図2)。おそらく、この分割阻害が原因となって、オッドペアードRNAi胚では鋏角と触肢が発生しなかった。ヘッジホッグのストライプよりもさらに前方では、分子マーカーの正常な発現が観察されたことから、オッドペアードの働きは鋏角と触肢の発生に特異的なものであることが示唆された。また、オッドペアードRNAi胚ではオッドペアード遺伝子自身の転写が大きく影響を受けて正常よりも広い領域で活発化していることから、負のフィードバック制御が働いていることが考えられた。
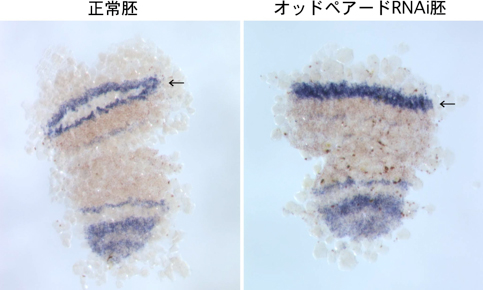 |
図2 オッドペアードRNAi胚の表現型
矢印で示しているように、頭胸部前方領域のヘッジホッグのストライプの分割が起こらない
オッドペアードはもともとショウジョウバエでペアールール遺伝子として同定された遺伝子である。この遺伝子はZnフィンガー型の転写因子をコードしており、ヘッジホッグシグナル経路の構成因子のキュービタスインタラプタスともフィンガー領域が類似している。脊椎動物では、オッドペアードの相同分子(ジック)とキュービタスインタラプタスの相同分子(グリ)がフィンガー領域を介して結合するという報告もある。クモの体節形成において、オッドペアードによる負のフィードバック制御がヘッジホッグシグナル経路と密接に関わりながら働いている可能性も十分にある。ヘッジホッグシグナル経路がダイナミックにパターンを生み出す基本的仕組みを解明するためにも、オオヒメグモの頭胸部における体節形成の解析を進めたい。
おわりに
私たちのオオヒメグモ初期胚を用いた研究は、細胞間シグナル経路の重要性を具体的に明らかにしている。ショウジョウバエ初期胚では細胞間シグナルはそれぞれ、比較的局所においてしか働いていない。そのために、パターン形成に必要な情報を細胞から細胞へどのように遠距離に伝達し、それらを統合するかという問題に取り組むには、ショウジョウバエは必ずしも適した材料ではない。それに対して、オオヒメグモ胚はその問題に取り組むことを可能にしてくれる絶好の材料である。今後この新しいモデル動物を用いて、動物の胚発生に関わる未知の分子機構を明らかにしたいと考えている。