年度別活動報告
年度別活動報告書:2015年度
脊索動物と節足動物の共通祖先を理解する 2−1 オオヒメグモの胚発生を支配する分子メカニズムの解明
小田広樹(主任研究員) 秋山-小田康子(特別研究員)
岩﨑佐和(奨励研究員) 野田彰子(研究補助員)
はじめに
多細胞動物は形態や発生様式に基づいて「門」と呼ばれる高次分類群(例えば、ヒトを含む脊索動物門や昆虫を含む節足動物門)に区分されているが、それぞれの分類群を特徴付ける形態が進化してきた過程やその進化の仕組みを理解することは簡単ではない。その大きな要因のひとつは、分類群を特徴づける、機能性の高い進化的に安定な形態であっても、その形態を形作るための発生過程やゲノム情報(遺伝子発現制御機構や遺伝子ネットワークなどを含む)は進化の過程で多様に変化しうることにある。
例えば、体の軸に沿った反復構造の存在は、ヒトなどの脊椎動物を含む脊索動物門と昆虫やクモを含む節足動物門に共通して見られる特徴であるが、その反復パターンを形作る遺伝的仕組みに両者が同一起源であることを示す確かな証拠は見出されていない。しかし、だからと言ってその類似の形態的特徴が異なる起源から別々に進化したと断言することはできない。なぜならば、共通祖先において存在した反復パターンを形成する仕組みが、発生プログラムのアウトプットとなる反復パターンを保持したまま、現存種のゲノムの比較では簡単に追跡できないほど大きな変化を経て多様化したかもしれないからである。このような可能性を考えた場合、形態で分類されるそれぞれの動物門での遺伝的仕組みの多様性の程度を大局的に把握することが動物門間の進化的関係を理解する上で重要である。さらに、形態進化とゲノム進化の基本原理を理解するという観点から言えば、特定の形態やパターンを発生プログラムのアウトプットとして出し続けながらそのプログラムがどのように進化し、多様化しうるかを、システム生物学や数理生物学を取り入れて解析することも重要となる。
私たちはこれまで独自のモデル生物種として、節足動物門鋏角類オオヒメグモ (Parasteatoda tepidariorum) を用いて、節足動物の基本的体制(ボディープラン)に関わる発生メカニズムの多様性を明らかにしてきた。オオヒメグモ胚では、ヘッジホッグシグナルとディーピーピーシグナルを介した細胞間のコミュニケーションが直交した2つのからだの軸(前後軸と背腹軸)の形成に主要な役割を果たす1),2)。ヘッジホッグシグナルの初期胚での役割の重要性は、節足動物の進化において発生プログラムが大きな変化を受けたことを象徴的に示す。オオヒメグモ初期胚で分泌タンパク質のヘッジホッグが果たす役割と同様の役割をショウジョウバエでは転写因子のビコイドが果たす3)。さらに、ヘッジホッグやビコイドの支配下で起こる縞パターン形成(体節形成)にもオオヒメグモとショウジョウバエで違いがある。つまり、発生プログラムのアウトプットとして縞パターンは両者で保存されているが、その縞パターンの形成過程は大きく異なる4)。クモを起点とした比較研究は、ボディープランの保守的進化の中で発生過程や発生メカニズムがどのように多様化したのか、また、どうして多様化できたのかを理解することに貢献しうる。これは、遺伝的発生システムの進化能の問題であり、理論研究とも連携すべき研究課題である5)。
本年度私たちは、オオヒメグモのゲノムに関する情報を効率的に利用するための枠組みを整備するとともに、オオヒメグモ初期胚の軸形成の仕組みをゲノムワイドに解析するために、次世代シーケンサーの利用を強力に押し進めた。昨年度より行っているヘッジホッグシグナル制御下の遺伝子ネットワークの解析に加え、オオヒメグモ初期胚の局所から取得した微量サンプルに基づく遺伝子発現解析を新たに試みた。また、昨年度ハエトリグモを用いた移植による重複胚の作出を報告したが、研究の発展性を考えて、本年度オオヒメグモ胚を用いて移植でなく部分的除去で重複胚の作出を行った。まとめる段階にある研究を含めて、本年度の研究活動を報告する。
結果と考察
(1)オオヒメグモのDNA塩基配列データベースの構築と公開
2013年度の報告書に、オオヒメグモDNA塩基配列データベースPt_spiderBASEの構築と公開について報告したが、レンタルサーバのみで運営していたためにBLASTによる検索機能が利用できなかったが、今年度、BRH独自にWebサーバを立ち上げ、BLAST検索を利用できるようにした。
さらに、BRHの研究で取得したデータのみから構成されるPt_spiderBASEに加えて、ベイラー医科大学で得られたオオヒメグモのゲノム配列のデータ6)と国際的オオヒメグモ研究者コミュニティ7)で行ったトランスクリプト予測のデータを取り込んだデータベース(Pt_spider Genome & Transcripts Data BASE)を構築した。現在まだオオヒメグモゲノムの論文が未発表であるためにこのデータベースのデータを使った研究成果の公表には制限があるが、論文作成に向けて研究者コミュニティ間でデータが共有化されている。私たちが作製したデータベースもドメインが取得され次第、世界のオオヒメグモ研究者と共有化を図る予定である。
これら2つのデータベースへは新しく立ち上げたBRH Webサーバのトップページからアクセスできる(図1 )。
(2)発生段階を追った遺伝子発現プロファイルの取得とデータベース化
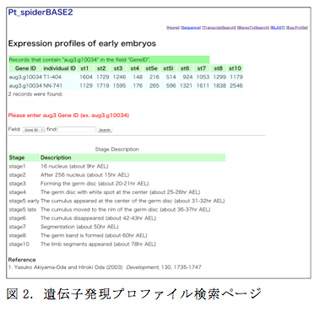
オオヒメグモ初期胚の体軸形成メカニズムを解析する基盤を作るため、RNA-seqを用いて初期胚の時間軸に沿った遺伝子発現プロファイルを取得した。2匹の雌個体から独立に集めたステージ1から10までの胚から、2シリーズのライブラリを作製した。次世代シーケンサーMiSeqを用いて150bpのシングルリードで各サンプルにつき1000万リード程度の塩基配列を取得した。配列情報はBLATアルゴリズムを用いてゲノムにマッピングし、それらの結果をHTSeq-countプログラムを用いて遺伝子発現量としてカウントした。さらに、私たちはこれらの情報をデータベース化した。 前述のPt_Spider Genome & Transcripts Data BASE中のExpression profile searchページでは、Gene IDを入力し検索をかけるとステージ1から10までの遺伝子発現量が表示される仕組みになっている(図2)。これにより、世界中のクモ研究者コミュニティでの情報共有を目指す。
(3)クモ初期胚でヘッジホッグシグナルに制御される遺伝子の同定と発現解析
ヘッジホッグシグナルは動物の発生において、脊椎動物の神経管や肢芽、ショウジョウバエの体節などの様々な組織の極性形成にはたらく。オオヒメグモ初期胚では胚全体の極性形成を支配し、直交する2つの体軸の形成を制御することを私たちはこれまでに明らかにしてきた。この結果は、発生の比較的後期に起こる現象だけではなく、最も初期の現象である体軸形成においてもヘッジホッグシグナルが重要な役割を果たすことを示している。初期胚が非常にシンプルな構造であることも考え合わせると、ゲノムワイドな探索によるクモ胚の極性形成・体軸形成に関わる遺伝子ネットワークの全貌の解明が、動物の体軸形成の進化やヘッジホッグシグナルの役割の進化に関する新たな理解と、複雑なネットワークを作ることが知られているヘッジホッグシグナルの全体像の解明につながると期待できる。
私たちはparental RNA interference (pRNAi)とMiSeqを利用したRNA sequencing (RNA-seq)を組み合わせた方法で、ヘッジホッグシグナルに制御される遺伝子の探索を行っている。昨年度までに、ヘッジホッグに対するpRNAiを行い、続いて行ったRNA-seqのデータをオオヒメグモゲノムにマッピングすることで、pRNAiの前後で発現量に変動の見られる遺伝子をゲノムワイドに同定しうることを確認した。今年度はまず、ドイツのグループが予測したオオヒメグモゲノム上の遺伝子構造の情報が利用できるようになり、これが有用であることが確認できたので、この情報を活用した新たな解析方法を確立した。続いて、ヘッジホッグシグナルの負の制御因子であるパッチトと、実験のコントロールとしてクラゲの蛍光タンパク質gfpに対するpRNAi & RNA-seqのデータも合わせて解析し、ヘッジホッグとパッチトのどちらのpRNAiでも発現が変動する87の候補遺伝子を同定した。ここにはヘッジホッグシグナルで制御されることが既知の遺伝子や以前に行ったマイクロアレイで同定された遺伝子が含まれており、方法が確かに動いていることが示唆された。Gene Ontology解析の結果、脊椎動物の肢芽形成に関わる遺伝子が多く含まれることが分かった。
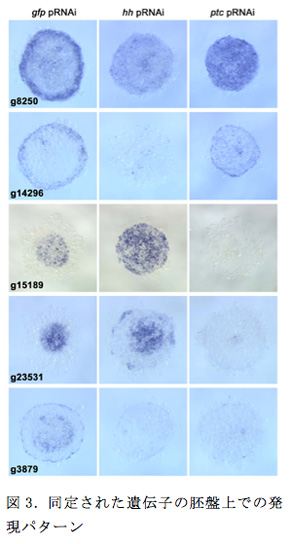
これら87遺伝子のうち19は以前にクローニングを行っており、37はESTに含まれていたためクローンをおこし、残り31をPCRでクローニングした後、全ての遺伝子のプローブを作製し、in situハイブリダイゼーションで発現を解析した。2つの体軸が直交するステージ5と続くステージ6(またはどちらか)で、46遺伝子の発現が領域特異的に検出された。ヘッジホッグのpRNAiで発現が下がりパッチトで上がることがRNA-seqの結果から示唆された遺伝子の多くは、将来頭部を形成する胚盤の周縁領域で発現が見られ(e.g., g8250, g14296)、反対にヘッジホッグで上がりパッチトで下がる遺伝子の多くは将来後体部を形成する胚盤の中心領域で発現が見られた(e.g., g15189, g23531)。またヘッジホッグ、パッチトで共に発現が下がる遺伝子の多くは中間領域で発現が検出された(e.g., g3879)。7遺伝子が背側を誘導するCM細胞で、3遺伝子がCM細胞と接する胚盤上皮細胞で発現が確認された。まだ現在進行中であるが、これらの胚盤上の発現がヘッジホッグに依存していることをいくつかの遺伝子で確認している(図3)。
このようにヘッジホッグシグナルに応じて様々な遺伝子が胚盤の各領域で発現することが分かってきた。多数の遺伝子の発現の重なり合いを考えると、様々な異なる状態の細胞が胚盤の半径に沿って存在することになる。今後はさらに、ここで見つかってきた遺伝子が次にどのような現象を引き起こすのかを解析する予定である。ここで明らかになった結果をデータベースに反映させることも計画している。
(4)微量サンプルRNA-seqによる初期胚の領域特異的発現遺伝子の同定
オオヒメグモ初期胚の体軸形成を理解するためには、初期胚の遺伝子発現を空間的に把握する必要がある。そこで本年度は、初期胚の一部の細胞を採取し、RNA-Seqによりゲノムワイドな遺伝子発現解析をおこなう手法の確立を行なった。具体的には、ガラスキャピラリーを用いてステージ5の胚盤中心に位置するクムルスあるいは縁の細胞を採取しmRNAを抽出。微量のmRNAサンプルからライブラリ合成を行なえるSMARTer Stranded RNA-Seq Kit(Takara Clontech社)を用いてライブラリを作製し、MiSeqのシーケンシングにより200〜500万リード程度の塩基配列を取得した。データ解析ではedgeR等のプログラムを用いて中心と縁で発現に差のある遺伝子を114個抽出した。さらにステージ4とステージ3でも同様の手法で空間的に遺伝子発現に差がある遺伝子を探索している。
(5)オオヒメグモの胚操作による部分重複胚の作出
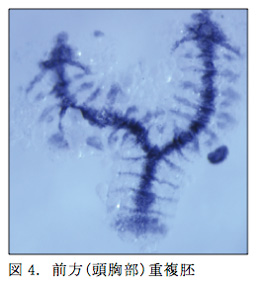
鋏角類のクモの初期胚は高い調節能を持ち、重複胚を形成することが古くから知られていた。Holmはイナズマクサグモ(Agelena labyrinthica)の卵を用いて、実験的にクムルスを別の場所に移植することで重複胚を作成している8)。しかし近年クモで実験的な重複胚の報告はなく、調節性の高い体軸形成メカニズムはまだ明らかになっていない。本年度は、オオヒメグモ胚のステージ 5 初期において、胚の一部を切除することで前方(頭胸部)重複胚を約20-30%の確率で作出できることを発見した(図4)。今後、これら前方重複胚の形態構造を確認するとともに、切除によって撹乱された胚において体軸がどのように決定されるのかを明らかにしていく予定である。
(6)反復性を示す縞パターン形成ダイナミクスの定量解析
オオヒメグモ胚の予定頭部領域と予定後体部領域で反復性を示す、2つの異なる縞パターン形成動態がヘッジホッグの遺伝子発現パターンの変化として観察される。2013年度、2014年度の報告書で示したように、タイムシリーズの固定胚を用いて縞パターン形成動態の定量解析を行ってきたが、本年度、同じデータセットを基に胚の形状指標S(2014年度報告書参照)と固定時間Tの関係を用いて反復性の縞パターン形成動態の1サイクルにかかった時間を見積もった。予定頭部領域では既存の縞が前後に非対称に2回分裂するが、分裂ピーク間の距離のSに対する変化速度を基に分裂開始点を定義すると、1回目の分裂と2回目の分裂の開始点のSの差は約1.3で、約5時間に相当した。一方、予定後体部領域では繰り返し新たに点状の発現領域が出現し、縞状のパターンに発展するが、後端部領域に設定した測定ポイントにおけるシグナル強度のSに対するプロットから周期性が観測され、その周期はSの差で約1.2〜1.3で、時間に換算すると約4.5〜5時間であった。
さらに、予定後体部領域において観察されるヘッジホッグの発現領域の形やサイズの変化がどの程度の数の細胞で引き起こされているかを具体的に見積もるために、異なるS値を示す7個の胚に関して、発現領域ごとに細胞数をカウントした。その結果、Sが1増加する間(約4時間)にヘッジホッグを発現する細胞が新たに現れ、その数を300以上にまで増やすことが想定された。細胞標識の解析では、胚盤期のステージ5初期から後体部第二体節形成期までの間(約20時間以上)に3回程度までの分裂しか観察されていないことを考えると、胚帯後端部領域でのヘッジホッグの発現領域の拡大を細胞分裂だけで説明することはできない。また、同じ細胞標識のデータに基づくと、大量の細胞が他の領域からまたは胚内部から胚帯後端部表層へ移入してくることも考えにくい。以上の状況を考え合わせると、予定後体部領域でのヘッジホッグの遺伝子発現パターンのダイナミックな変化は個々の細胞での発現動態を反映している可能性が高い。個々の細胞レベルでの遺伝子発現動態を想定すれば、点パターンの出現から、点パターンの拡大、ドーナツ形を経て縞パターンへの発展のサイクルは同調的な発現振動で容易に説明しうる。遺伝子発現の振動現象を直接観察することはできていないが、オオヒメグモ胚の予定後体部領域では同調的振動による縞パターン形成機構の存在が強く示唆される。
おわりに
本年度は、BRH独自のWebサーバの構築により、BRHから学術情報を世界に発信する環境が整った。多量に創出されるデータを研究室内だけでなく、研究室外の研究者コミュニティと効率的に共有することは研究分野の健全な発展のために欠かせない。日々の活動で新たな知識の発見に注力することはもちろん大事であるが、長期的にみて、知識の発見を効率化することも重要であると認識しており、それぞれのテーマの研究で創出されたデータを独自にデータベースにまとめて公開していきたい。
初期胚の局所から得た細胞である程度の精度のRNA-seq解析ができたことは研究の幅を広げる大きな成果であった。オオヒメグモ初期胚の極性形成のプロセスを解析するための新たな方法論を得ただけでなく、eRNAiとの組み合わせでRNA-seq解析をする道が開けた。また、オオヒメグモ胚の胚盤部分除去により、重複胚を作ることに成功したことは、オオヒメグモ胚の発生調整能の高さを示すものであり、オオヒメグモを実験材料として使用することに対する強力な根拠となりうる。