年度別活動報告
年度別活動報告書:2004年度
脊索動物と節足動物の共通祖先を理解する 2-2 クモの体軸形成と中胚葉形成の解析
小田広樹(研究員)
秋山-小田康子(派遣研究員)
野田彰子(研究補助員)
入江雅美(派遣実験補助員)
はじめに
近年の分子遺伝学的な研究は、脊椎動物とショウジョウバエが個体発生の似たような場面で相同な遺伝子を使っていることを明らかにした9)。例えば、BMP/dppとchordin/sogの活性で引き起こされる背腹軸形成、hairyで制御される体節形成、Hoxクラスターで制御される前後軸に沿った体の領域化、Pax6/eyelessで制御される眼の形成、Nk2.5/tinmanで制御される心臓の形成などが挙げられる。これらの発見に基づいてUrbilateria仮説が提唱され、”すべて”の左右相称動物の共通祖先はそれらの発生形質を持った比較的複雑な動物であったと考えられるようになった。この仮説は、一般的に信じられている左右相称動物の系統関係を取り込んで、脊椎動物とショウジョウバエの共通祖先とすべての左右相称動物の共通祖先が一致することを前提にしている。しかし本来は、その系統関係が証明されない限り、脊椎動物とショウジョウバエで見られる類似性の原因がすべての左右相称動物の共通祖先の特徴を引き継いでいるためなのか、それともショウジョウバエと脊椎動物が他の動物に比べて近縁であるためなのかを区別することはできない(もちろん、収斂進化による可能性も残されている)。この議論は左右相称動物の初期進化だけでなく、脊椎動物の起源を理解する上で極めて重要である。しかし、この議論を発展的に展開させるには節足動物門の中の多様性に関して私たちはあまりにも無知である。節足動物はカンブリア紀の最も初期から豊富な化石記録があり、それらの形態的な特徴はUrbilateria仮説が想定するものと似ている。しかし、その形態を作り出すための発生メカニズムは同じ節足動物門の中でも長い進化の過程で大きく多様化したはずである。初期の節足動物の発生メカニズムがどのようなものであったのかを理解することは、節足動物門と他の動物門との関係性を理解する上で必要不可欠である。
本研究では、節足動物の祖先的な発生メカニズムを理解するために、鋏角類のクモを使ってショウジョウバエとの比較解析を行っている10)。特に、動物門を越えて比較しうる発生現象の重要性を考え、胚葉形成と体軸形成に集中して解析している。本年度は、オオヒメグモにおけるparental RNA干渉法(pRNAi法) の確立に成功し、遺伝子の機能解析に道が開けた。背腹軸形成の解析では、そのメカニズムにハエとクモの間で大きな違いがあることを明らかになった。また、中胚葉運命を決定するメカニズムにも大きな違いがあることがわかってきているのでそれも合わせて報告する。
結果と考察
昨年度までは遺伝子発現に基づいた比較解析を行ってきたが、本年度は遺伝子機能を解析するための方法開発を行った。線虫やトリボリウム(甲虫) で報告されているparental RNA干渉法 (pRNAi法)をオオヒメグモに対してその効果を調べた。成熟した雌グモにin vitroで合成したAt.sogの二本鎖RNAを1回あたり2~4_gを注入し、そのクモが産んだ卵の発生を解析した(コントロールとして別の雌グモにGFPの二本鎖RNAを注入した)。1回の注入では効果はほとんど見られなかったが、3、4日おきに3~4回繰り返すことにより100%の胚で特異的な異常が観察されるようになった (図3、後述)。その効果は注入回数が多い程強く見られ、かつ長期間(計4回の注入で50日以上)持続した。RT-PCR法およびin situハイブリダイゼーション法によってAt.sog mRNAの発現が特異的に抑制されていることも確認した。さらに、At.dppに関して同じ実験を行ったところAt.sogとは異なる表現型が得られた。これらのことから、オオヒメグモにおいてもpRNAi法が遺伝子機能を解析するための方法として使えることが示された。
At.sog RNAi胚では胚帯形成までは正常に進んだが、胚帯内での背腹パターンに明らかな異常が生じた。正常胚では肢芽が頭胸部の各体節に2つずつ、最も背側の領域に分かれて形成されるが、At.sog RNAi胚では各体節でひとつの大きな尾芽が形成された。分子マーカーの発現で表現型を解析したところ、At.sog RNAi胚では腹側正中線と神経性外胚葉が形成されておらず、胚帯の細胞すべてが背側の運命をとってしまったと考えられた。このことは、At.sogが腹側正中線領域と神経性外胚葉領域の特異化に必要であることを示唆する。
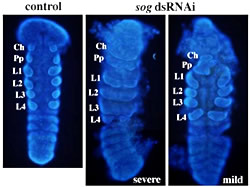 |
図3:pRNAiによってAt.sogの発現を抑制した胚の発生異常
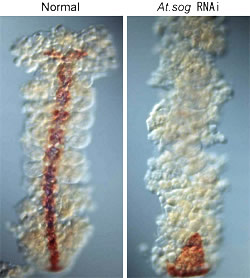 |
図4:正常胚とAt.sog RNAi胚をAt.sim(青、腹側正中線のマーカー)とAt.sog(茶)のプローブで二重染色したもの
一方、ショウジョウバエのsog突然変異体では、腹側正中線の形成に異常は現れず、神経も細胞の数が多少減るものの、クモのAt.sog RNAi胚ほど大きな異常としては現れない。ここで注目すべきは、sogの機能がクモでは腹側正中線の形成に必須であるのに対し、ハエではそうではないことである。この違いは、節足動物の腹側正中線と脊椎動物の脊索が相同である可能性を考えるとおもしろい。sogの脊椎動物相同分子はchordinであるが、この分子は脊索の形成及びこの組織を中心とする背腹軸の形成に極めて重要な役割を果たすことが知られている。脊椎動物胚の予定脊索領域は体軸を誘導する活性を持つが、クモの腹側正中線領域が背腹のパターンを誘導する活性(例えば、神経を誘導する活性)を持つかを検証することは重要であり、今後の課題である。さらに、節足動物の腹側正中線が外胚葉であるのに対し、脊椎動物の脊索が中胚葉であることを考えると、それぞれの胚葉の進化を理解する努力も必要である。
(2)オオヒメグモの中胚葉形成に関する解析
昨年度報告したように、ショウジョウバエのtwistのクモ相同遺伝子At.twistは中胚葉細胞と思われる細胞で発現しており、中胚葉の発生を解析するための貴重な分子マーカーとなる11)。本年度は、オオヒメグモの中胚葉細胞がいつ、どこから、どのように誕生するかを突き止めるために、初期胚の発生ステージを厳密に決め、At.twistの初期の発現を詳細に解析した。その結果、ステージ5の後期(クムルスが胚盤の縁付近に着いた時)に胚盤の縁でAt.twistを発現している細胞が見られるようになった。ステージ6に入ると(クムルスが胚盤の縁付近に到着後、胚盤細胞の再配列が始まると)、胚盤の周縁領域の他に、胚盤の中心領域でもAt.twistを発現している細胞が見られるようになり、発生が進むにつれてAt.twistを発現している細胞の数は急速に増えた。At.twistの発現解析に加え、βカテニンの抗体やファロイジンによる染色及びDNA染色の解析から、ステージ6の初期からステージ7の初期にかけて胚盤の中心領域または形成されつつある尾葉の周辺領域からAt.twistの発現を始めた細胞が胚内部に陥
入することが示唆された。
一方、理化学研究所の阿形清和博士らとの共同研究で行ったEST解析で得られた、ショウジョウバエのDeltaのクモ相同遺伝子At.Deltaの発現を解析したところ、胚盤の中心領域でAt.twistと似た発現パターンが観察されたので両者の発現を二重染色によって詳しく比較した。その結果、一部の細胞でAt.Deltaの発現がAt.twist発現と一致ししていることが分かり、さらに異なるステージの発現パターンの比較から、At.Deltaを発現し始めた細胞(At.Deltaの発現は一時的)がその後At.twistを発現するようになることが考えられた。これらの観察結果は、ショウジョウバエと違ってオオヒメグモでは中胚葉の運命決定にDelta-Notchのシグナルが関与している可能性を示唆する。また、ショウジョウバエでは母性転写因子であるDorsalが直接、twistの転写を活性化しているのに対し、オオヒメグモではtwistの転写制御に細胞間相互作用が介在してる可能性がある。節足動物の進化の過程で中胚葉の運命を決定するメカニズムに大きな変化があったことが示唆された。
おわりに
本研究によって明らかにされつつあるクモの体軸形成と中胚葉形成のメカニズムは、対応するショウジョウバエのメカニズムとは、細胞間相互作用に高い比重が置かれている点で異なっていると考えられる。このようなクモとハエの相違点の発見は、節足動物門内で起こった発生システムの変化プロセスを解き明かすための重要な手がかりを提供すると同時に、節足動物の祖先的な発生システムに関して理解を深めさせてくれる。冒頭にも述べたが、私たちの研究の一番の目的は動物門を越えた比較を可能にすることである。それが実際に可能かどうかはわからないが、もし可能であるならばそれぞれの動物門の祖先を追究した先には動物門の間に共通点が見えてくるはずだと期待している。
参考文献
1) Philippe, H., Chenuil, A., and Adoutte, A. (1994) Can the Cambrian explosion be inferred through molecular phylogeny? Development Suppl.: 15-25.
2) Abouheif, E., Zardoya, R., and Meyer, A. (1998) Limitations of metazoan 18S rRNA sequence data: implications for reconstructing a phylogeny of the animal kingdom and inferring the reality of the Cambrian explosion. J. Mol. Evol. 47: 394-405.
3) Rokas, A., and Holland, P. W. H. (2000) Rare genomic changes as a tool for phylogenetics. Trends Ecol. Evol. 15: 454-459.
4) Oda, H., Wada, H., Tagawa, K., Akiyama-Oda, Y., Satoh, N., Humphreys, T., Zhang, S., and Tsukita, S. (2002) A novel amphioxus cadherin that localizes to epithelial adherens junctions has an unusual domain organization with implications for chordate phylogeny. Evolution & Development 4, 426-434.
5) Oda, H., Akiyama-Oda, Y. and Zhang, S. (2004) Two classic-cadherin related molecules with no cadherin extracellular repeats in the cephalochordate amphioxus: distinct adhesive specificities and possible involvement in the development of multicell-layered structures. J. Cell Sci. 117, 2757-2767.
6) Wada, H., and Satoh, N. (1994) Details of the evolutionary history from invertebrates to vertebrates, as deduced from the sequences of 18S rDNA. Proc. Natl. Acad. Sci. USA 91: 1801-1804.
7) Turbeville, J. M., Schulz, J. R., and Raff, R. A. (1994) Deuterostome phylogeny and the sister group of the chordates: evidence from molecules and morphology. Mol. Biol. Evol. 11: 648-655.
8) Jeffery, W. R., Strickler, A. G., and Yamamoto, Y. 2004. Migratory neural crest-like cells form body pigmentation in a urochordate embryo. Nature 431: 696-699.
9) Carroll, S. B., Grenier, J. K., and Weatherbee, S. D. (2001) From DNA to diversity. Blackwell Science.
10) Akiyama-Oda, Y. and Oda, H. (2003) Early patterning of the spider embryo: A cluster of mesenchymal cells at the cumulus produces Dpp signals received by germ disc epithelial cells. Development 130, 1735-1747.
11) Yamazaki, K., Akiyama-Oda, Y. and Oda, H. (2005) Expression patterns of a twist-related gene in embryos of the spider Achaearanea tepidariorum reveal divergent aspects of mesoderm development in the fly and spider. Zool. Sci. 22 (in press).