年度別活動報告
年度別活動報告書:2004年度
脊索動物と節足動物の共通祖先を理解する 2-1 左右相称動物におけるアドヘレンスジャンクションの多様化と分子シナポモルフィーの同定
小田広樹(研究員)
秋山-小田康子(派遣研究員)
野田彰子(研究補助員)
入江雅美(派遣実験補助員)
はじめに
生物の系統関係を推定する方法として、相同なDNA配列に起こった塩基置換に基づいて確率論的に推定する方法が最も一般的に使われているが、この方法だけでは系統間や分子内の部位間による置換速度のばらつきなどが主な原因となって系統樹の深いところの分岐パターンを必ずしも正確に推定することはできない1), 2)。そこで近年は、塩基置換に基づく推定方法を補完するためにも、ゲノム中に見られる相同形質において、ある動物群だけに共有されている派生的な状態(シナポモルフィー,synapomorphy)を見つけだすことによって共通子孫を特定しようという試みがなされている3)。しかし実際には、そのような理想的な形質をゲノムの中に同定することは極めて難しい。
私たちは昨年までの研究で、多細胞動物の上皮においてアドヘレンスジャンクション(adherens junction)の形成を担う接着分子カドヘリンのドメインの構成に複数の状態があることを明らかにし4), 5)、さらにドメインの比較解析とそれぞれの構造状態の左右相称動物における分布パターンから、祖先的状態とシナポモルフィーと見なせる派生的状態があることを可能性として示した。多細胞動物にとって普遍的かつ重要な相同構造の中に分子的なシナポモルフィーが同定できることは、動物の系統分類を考える上で極めて大きな意義を持つ。本年度は、さらに詳しいドメインの比較解析によってカドヘリンの構造状態のシフトを明確に示唆するデータを新たに得たので報告する。さらに、得られたデータに基づいて細胞間結合構造の多様化と動物の系統発生との関係性を考察したのでそれも合わせて報告する。
結果と考察
私たちが昨年度までに同定した左右相称動物のカドヘリン及びすでに公共のDNAデータベースに登録されているカドヘリンのドメイン構成は、少なくとも8つのタイプ (A1、A2、E1、E2、H、C、V1, V2) に分類される。例えば、A1のドメイン構成はハエやコオロギ、トビムシなどの六脚類に加え、鰓脚類のアルテミアで見つかっており、V1のドメイン構成は多数の脊椎動物に加え、尾索類のホヤで見つかっている。同じ構造状態が綱などの比較的高次の分類群を越えて保存されていながら、系統的にさらに遠く離れていると思われる動物間では構造状態が大きく異なっているという事実は、カドヘリンのドメイン構成の多様性が動物の初期進化における遺伝的な変化を反映している可能性を示していると考えられる。
そこで、カドヘリンのドメイン構成の多様性がどのように形成されたかを調べるために、それぞれのカドヘリンのアミノ酸配列をドメインごとに分割し、それらを別のカドヘリンのアミノ酸配列全体に対してBLASTPにかけ、得られたE-valueをマトリックス上にプロットした。この解析により、2つのカドヘリンの間で相同なドメインを検出することができる。このような解析を様々なカドヘリンの組み合わせで行い、カドヘリンのドメイン構成がどのように進化したかを検討した。
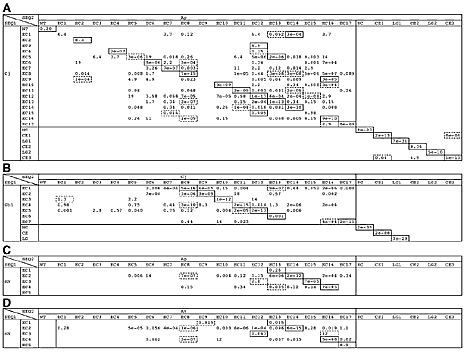 |
図1:BLASTを用いたカドヘリンのドメイン構成の比較解析
(A) エビのカドヘリン (Cj) とヒトデのカドヘリン (Ap) の比較。(B) コオロギのカドヘリン (Gb1) とエビのカドヘリン (Cj) の比較。(C) マウスのカドヘリン (mN) とヒトデのカドヘリン (Ap) の比較。(D) マウスのカドヘリン (mN) とクモのカドヘリン (At) の比較。アミノ酸配列がお互いによく似ているドメインが四角で示されており、どのドメインとどのドメインが相同であるかを推定することができる。エビとヒトデのドメイン構成を祖先的であると考え、コオロギとマウスの異なるドメイン構成が祖先的状態からのドメインの欠失によって生じたと考えると分かりやすい。
その結果、A2, E2, V2のドメイン構成はお互いによく似ているが、この類似は進化的保存によるものであり、収斂進化による可能性は低いと考えられた(図1A)。
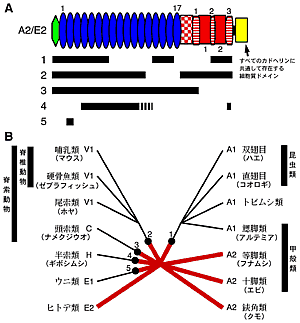 |
図2:カドヘリンの祖先的構造状態(A2/E2)から派生的構造状態が生じた時に欠失した領域(A)とその欠失変化の系統樹へのマッピング(B)
A2,E2,V2のカドヘリンは細胞質ドメインのアミノ酸配列における置換に基づいた系統解析では必ずしも近縁性を示さないので、A2/E2が左右相称動物の祖先的状態であると考えるのが最も考えやすい。そこでA2/E2
を祖先的状態であるとすると、図1Bの結果はA1が3つの領域の欠失によって誕生したこと、図1C, Dの結果はV1がA1とは異なる欠失変化で誕生したことを示唆する。またA1, V1と同じように、C、E1、Hの誕生もA2/E2からの欠失変化で説明できる。図2Aにそれぞれの派生的状態の誕生において欠失したと考えられる領域をまとめた。この図から明らかなように、少なくともA2/E2→A1、A2/E2→V1、A2/E2→Cの変化は独立的に起こったと考えられる。この独立性は、すべてのクラシックカドヘリンに共通して存在する細胞質ドメインにおけるアミノ酸配列の置換パターンとも矛盾しない(つまり、図2Bと同じトポロジーの系統樹が得られる)。そしてさらに、A1, V1, C, E1, Hの構造状態が左右相称動物において、お互いに排他的に分布していることとも辻褄が合う。これらの一連のデータは、祖先的状態(A2/E2)からシフトしたカドヘリンの構造状態(A1,V1,Cなど)がシナポモルフィーとしての条件を満たす可能性があることを示している。この可能性をより厳密に検証するには、カドヘリンの構造に関する情報を網羅的に集め、矛盾が生じないことを示す必要があるであろう。つまり、カドヘリンの構造状態のシフトが進化の過程で稀にしか起こらなかったことと、派生的であると考えている構造状態が確かに、お互いに排他的に分布していることを示さなければならない。
私たちの仮説的提案は、カドヘリンの生理機能を考えると興味深い。A1,V1,Cなどのカドヘリンの派生的構造状態は、それぞれの動物において上皮のアドヘレンスジャンクションの形成を担う。カドヘリンは特異性の高いホモフィリックな接着活性によって多細胞体の形成に寄与し、特に上皮のカドヘリンは個体と外界を分ける上皮細胞層を形作る。ある動物集団のある個体の上皮においてカドヘリンの構造に変化が起こり、異なる接着活性を持つカドヘリン分子が形成されたと想定すると、これによって祖先的な細胞集合体とは認識し合うことができない新たな細胞集合体が生まれた可能性が考えられる。カドヘリンの大胆な構造変化が、異なる細胞集合体の形成及びその集合体の独立的な進化に貢献したかもしれない。
もし私たちの仮説的提案が正しければ、動物の系統関係を見直す必要があるであろう。A1の構造状態は六脚類と鰓脚類が近縁であることを支持する(図2B)。古典的には、鰓脚類は甲殻類の中でも原始的なグループに分類されている。しかし、私たちのデータは十脚類や等脚類を原始的なグループに分類し、鰓脚類と六脚類がその原始的なグループから派生した同一のクレードに入っていると考えると理解しやすい。現在、六脚類が甲殻類の中から誕生したことに関しては定説になりつつあるが、六脚類が甲殻類の中のどこに起源を持つかということに関してはまだまだ根拠が乏しい。私たちの結果はこの問題に対して新しい洞察を与えている。
V1の構造状態は脊椎動物と尾索動物が近縁であることを支持する(図2B)。これまで形態やDNAの塩基置換に基づく系統解析から脊椎動物に最も近縁な無脊椎動物は頭索動物であると考えられてきたが6), 7)、私たちの仮説はその考えを支持しない。脊椎動物のシナポモルフィーであると考えられている最も重要な形態形質のひとつに神経冠細胞があるが、つい最近の研究で尾索動物のホヤにも分子的に支持される神経冠細胞様の細胞が見つかった8)。一方、体節は脊椎動物と頭索動物を結び付ける重要な形態形質はであると考えられているものの、この考えに分子的な根拠は乏しい。Urbilateria仮説で言われているように左右相称動物の共通祖先が体節を持つ動物であったならば、尾索動物に体節がないという事実は体節性の二次的な喪失によって説明されうる。これらのことを考えれば、私たちが提案する脊索動物の系統関係は検討に値する。私たちの仮説を検証するには、より多方面からの分子的な根拠の積み重ねが必要であろう。
おわりに
本研究を通じて私たちは、動物の系統分類に関する新しい見方を提供できたと考えている。つまり、上皮の細胞間結合形成を担うカドヘリン分子の構造の多様性が左右相称動物の初期の系統分岐を反映しているかもしれないことを示した。これは直接的に証明できない以上あくまでも仮説でしかないが、様々な動物からのゲノム情報が蓄積すればある程度の検証は可能である。また、クモやエビを含む節足動物門の一部とヒトデを含む棘皮動物門の一部が左右相称動物の祖先的なグループとして位置付けられたことは、並行して進めているクモを使った発生進化の研究にとっても大きな収穫である。