年度別活動報告
年度別活動報告書:2005年度
脊索動物と節足動物の共通祖先を理解する 2−2 カドヘリンの構造と機能の関係の解析
小田広樹(研究員)
秋山-小田康子(派遣研究員)
野田彰子(研究補助員)
春田知洋(大阪大学大学院 M1)
はじめに
多細胞動物は細胞と細胞を連結するための特別な微細構造を持ち、それを巧みに操ることによって体の形を作り上げている。このような細胞間結合構造には何種類かのタイプがあるが、アドヘレンスジャンクションは最も普遍的に見られるタイプのひとつである。アドヘレンスジャンクションで細胞と細胞をくっつけている主要な分子はクラシックカドヘリン(以後、カドヘリンと呼ぶ)である。私たちはこれまでの研究で、カドヘリンの細胞外領域のドメイン構成は左右相称動物の間で多様であり、その多様性はヒトデやクモで共通に見られる状態からの独立的な短縮化で説明できることを見いだした 6)。短縮化されたあるひとつの状態は、ハエ、コオロギ、トビムシ、アルテミアで見られ、それとは別の状態が脊椎動物とホヤで、そして、さらに別の状態がナメクジウオで見られる。それぞれの状態は進化的に安定であるが、過去のある時点では大きな変化があったと考えられる。私たちは、そのようなカドヘリンの構造変化がカドヘリンの機能に変化をもたらしたかどうかを検討することによって、動物の形態進化とジャンクションの進化を関連づけたいと考えている。本年度はまず、昆虫型のカドヘリンの代表としてジョウジョウバエのカドヘリン(DEカドヘリン)を用いて、それぞれのドメインがそのカドヘリンの機能にどう関わっているかを調べた。
結果と考察
DEカドヘリンは、細胞外領域に7個のカドヘリンドメイン(EC1-7)と、無脊索ドメイン(NC)、システインリッチEGF様ドメイン(CE)、ラミニングロビュラードメイン(LG)で構成される原始クラシックカドヘリンドメイン(PCCD)複合体を持つ(図4)。
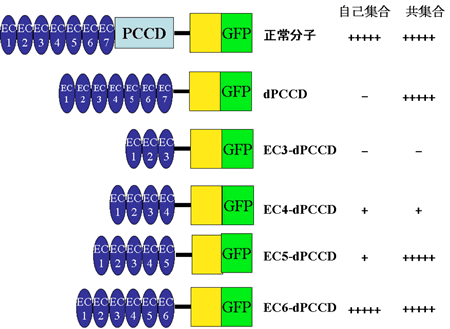 |
図4:作製した変異DEカドヘリン分子とその活性
PCCD複合体は、脊椎動物やホヤのカドヘリンには存在しない。これまでの解析でN末側のECドメインは接着に必須であることが分かっていたが、細胞膜貫通ドメイン(TM)に近い側のドメインが接着に関わっているのかどうかは不明であった。そこで、図4に示すように一連の欠失変異型分子を作製し、それぞれの分子の接着活性をショウジョウバエの培養細胞(S2細胞)に発現させ、集合実験を行うことによって調べた。集合実験では2種類の実験によって、変異分子を発現する細胞だけでどの程度の集合塊を作ることができるか(自己集合活性と呼ぶ)と、正常分子を発現する細胞と変異分子を発現する細胞を共集合させた時にできる集合塊に後者の細胞がどの程度入り込むことができるか(共集合活性と呼ぶ)を判定した。その結果、PCCD複合体だけを欠いた分子では共集合活性は見られたが、自己集合活性は見られなかった。にもかかわらず、PCCD複合体とEC7を欠いた分子では自己集合活性と共集合活性の両方が正常分子と同じ程度に見られた。このことは、EC7とPCCD複合体はS2細胞におけるカドヘリン接着に必ずしも必要ではないことを意味すると同時に、EC7からTMまでの領域の構造はその欠失のあり方によっては接着に対してネガティブな影響を与えうることを意味する。また、さらに欠失領域をさらに大きくした分子ではECドメインを少なくするほど集合活性が減少し、細胞外領域にEC1-EC3のみを持つ分子では自己集合活性も共集合活性も全く見られなかった。脊椎動物のカドヘリンではN末側の2つのECドメインだけでいくらかの接着活性を示すことが報告されているが 7) 、DEカドヘリンでは少なくともN末側の4つのECドメインは接着に必要であるようだ。
おわりに
EC7とPCCD複合体を欠いた分子でも培養細胞においては正常分子と同じレベルの接着活性を発揮することができる。それでは、EC7とPCCD複合体はカドヘリン機能にとって重要ではないのだろうか? 今後は、ショウジョウバエの個体発生においてそのような分子が十分に機能できるのかどうかを検討し、もし機能できないのであればその原因を追究したいと考えている。この解析によって、無脊索動物のカドヘリンに普遍的に存在し、脊椎動物やホヤのカドヘリンには存在しないPCCD複合体の機能的意義を理解することができるのではないかと期待している。
参考文献
1) Carroll, S. B., Grenier, J. K., and Weatherbee, S. D. (2001) From DNA to diversity. Blackwell Science.
2) Akiyama-Oda, Y. and Oda, H. (2003) Early patterning of the spider embryo: A cluster of mesenchymal cells at the cumulus produces Dpp signals received by germ disc epithelial cells. Development 130, 1735-1747.
3) De Robertis, E. M., Larra![]() n, J., Oelgeschl
n, J., Oelgeschl![]() er, M. and Wessely, O. (2000). The establishment of Spemann's organizer and patterning of the vertebrate embryo. Nat. Rev. Genet. 1, 171-181.
er, M. and Wessely, O. (2000). The establishment of Spemann's organizer and patterning of the vertebrate embryo. Nat. Rev. Genet. 1, 171-181.
4) Yamazaki, K., Akiyama-Oda, Y. and Oda, H. (2005) Expression patterns of a twist-related gene in embryos of the spider Achaearanea tepidariorum reveal divergent aspects of mesoderm development in the fly and spider. Zool. Sci. 22, 177-185.
5) Artananis-Tsakonas, S., Rand, M. and Lake, R. (1999) Notch signaling: cell fate control and signal transduction in development. Science 284, 770-776.
6) Oda, H., Tagawa, K. and Akiyama-Oda, Y. (2005) Diversification of epithelial adherens junctions with independent reductive changes in cadherin form: identification of potential molecular synapomorphies among bilaterians. Evol. Dev. 7, 376-389.
7) Shan, W., Yagita, Y., Wang, Z., Koch, A., Svenningsen, A.F., Gruzglin, E., Pedraza, L., Colman, D.R. (2004) The minimal essential unit for cadherin-mediated intercellular adhesion comprises extracellular domains 1 and 2. J Biol Chem. 279, 55914-23.