年度別活動報告
年度別活動報告書:2005年度
脊索動物と節足動物の共通祖先を理解する 2-1 オオヒメグモの体軸形成と尾葉形成の分子メカニズム
小田広樹(研究員)
秋山-小田康子(派遣研究員)
野田彰子(研究補助員)
春田知洋(大阪大学大学院 M1)
はじめに
近年のモデル動物を用いた分子発生学的研究は、脊椎動物とショウジョウバエが個体発生の類似した現象で相同な遺伝子を使っていることを明らかにしてきた 1) 。例えば、前後軸に沿ったパターン形成を制御する仕組みや背腹軸形成を制御する仕組みには脊椎動物とショウジョウバエでよく似た部分あることが分かっている。しかし、系統的に遠く離れた動物種間の比較だけでは、そのような類似がどのような経緯で生じてきたのかを客観性をもって議論することはできない。特に、動物門という枠組みを超えて議論するには注意を要する。なぜならば、同じ動物門の中でも大きな違いがあることに私たちはいくらか気付いているからである。脊索動物門と節足動物門における体節形成はそのよい例である。脊索動物門に分類される脊椎動物は体節を作るが、同じく脊索動物門に分類されている尾索動物(ホヤなど)は体節を全く作らない。一方、節足動物門でもショウジョウバエと他の多くの節足動物とでは体節の作り方が大きく異なる。異なる動物門を的確に関連づけるには、それぞれの動物門の中の保存性と多様性を分子レベルでしっかりと把握する必要がある。そこで私たちは、ショウジョウバエと同じ節足動物門に属しながら、ショウジョウバエから系統的に遠く離れているオオヒメグモを用いて、胚発生の仕組みを分子レベルで解析している。ハエとクモを比較することによって、節足動物門内での保存性や多様性に関する知見を効率良く得ることができるのではないかと考えている。
本研究では将来的に動物門を超えて比較することを目標としており、それを可能にするような発生現象に重点を置いている。本年度はオオヒメグモの体軸形成と尾部領域の発生に焦点を絞って解析を進めた。ショウジョウバエのdecapentaplegic (dpp ) とshort gastrulation (sog ) の相同遺伝子の機能解析によって、これらの遺伝子が放射相称から左右相称への転換に重要な役割を果たしていることを証明した。そして、ショウジョウバエのDeltaとNotchの相同遺伝子の機能解析によって、Delta-Notchのシグナルが体節形成の場となる尾部領域の特異化に必要であることを明らかにした。それぞれの遺伝子の役割をショウジョウバエとオオヒメグモで比較すると、ハエとクモの発生システムの違いが分かってきた。
結果と考察
(1)At-sog は胚帯内の腹側領域のパターン形成に必要である
私たちは昨年度までにショウジョウバエのshort gastrulation(sog ) とdecapentaplegic (dpp ) のオオヒメグモ相同遺伝子(At-sog とAt-dpp )をクローニングし、その発現パターンを明らかにした。そして、parental RNA干渉法(pRNAi)による遺伝子機能の抑制実験によって、At-sog が胚帯内のパターン形成に働いていることを示唆するデータが得られていた。本年度は、At-sog RNAi胚においてマーカー遺伝子の発現を詳細に調べることによって、At-sog は胚帯の腹側正中線領域とその両脇にできる神経領域の特異化に必須であることが明らかになった。ややマイルドな表現型を示すAt-sog RNAi胚ではAt-sog そのものの発現が所々に島状に検出されたが、このような胚ではAt-sog の発現領域を取り巻くように神経細胞が形成されていた(図1)。このことは、神経形成が胚帯内における位置よりも、At-sog の発現に支配されていることを示唆した。
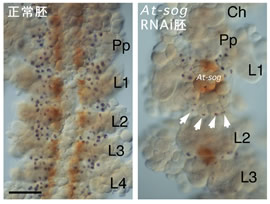 |
図1:At-sog RNAi胚のマイルドな表現型
残存するAt-sog の発現領域の周りに神経細胞が分化している(矢印)
(2)At-dppは放射相称から左右相称への転換に必要である
私たちはこれまでAt-dpp とリン酸化Madの発現解析から、オオヒメグモ胚では放射相称性を打ち破るように一方向に移動するクムルス間充織細胞(CM細胞)がDppシグナルを胚盤の上皮細胞層へ伝えることによって将来の背中側が決まるのではないかと推測していた 2) 。本年度はこのことを直接的に証明するために、pRNAiによってAt-dppの機能抑制実験を行った。その結果、At-dpp RNAi胚ではCM細胞の移動は正常に起こるものの、将来の背側にできるはずの胚外領域が形成されなかった。そして、各種のマーカー遺伝子で胚の相称性を調べたところ、At-dpp RNAi胚は発生が進んでも放射相称のままで、左右相称への転換が阻害されていることが分かった(図2)。また、At-dppRNAi胚ではAt-sog が尾葉を除く胚全体で強く発現していた。このことは、At-dppの活性がAt-sog の発現領域を腹側正中線領域に限局化するのに必要であることを示唆した。以上の結果をまとめると、At-dpp は放射相称から左右相称への転換に必要であり、At-dpp とAt-sog の相互作用によって腹側領域の特異化が行われているとが考えられる。
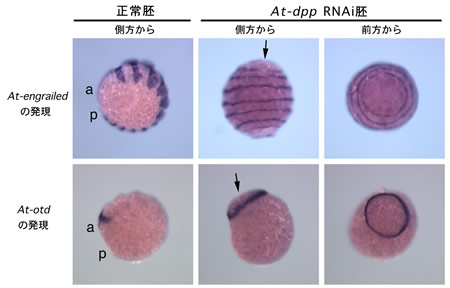 |
図2:At-dpp RNAi胚の表現型
At-dpp RNAi胚ではマーカー遺伝子の発現が環状に見られる
(3)ショウジョウバエとオオヒメグモにける腹側領域特異化の仕組みの違い
私たちの結果は、ショウジョウバエとオオヒメグモの腹側領域を特異化する仕組みに注目するとおもしろい。ショウジョウバエのsog 突然変異体では、腹側正中線の形成に異常は現れず、神経も細胞の数が多少減るものの、クモのAt.sog RNAi胚ほど大きな異常は現れない。つまり、sog の機能がクモでは腹側正中線と中枢神経の形成に必須であるのに対して、ハエではそうではない。ハエでは腹側領域の特異化が細胞間の相互作用よりもDorsalなどの転写因子に大きく依存している。ハエの多核性胞胚でDorsalは将来の背腹軸に沿って濃度勾配を作り、Dorsalの濃度が最も高い領域が腹側として特異化される。クモではDppシグナルによって背側がまず特異化され、その後、Dppシグナルを拮抗阻害するSogの活性によって背側からもっとも遠い領域が腹側として特異化されるようだ。このような細胞間相互作用によるシステムがクモにおける調節的な体軸形成を可能にしていると思われる。
クモの腹側領域特異化の仕組みは、脊椎動物の背側領域の特異化の仕組みと似ている。節足動物のDppとSogの相同分子は脊椎動物ではそれぞれBMP2/4とchordinとして知られているが、BMP2/4のシグナルをchordinなどで拮抗阻害することによって脊椎動物の背側領域が特異化される 3) 。このクモと脊椎動物の類似は、節足動物の腹側正中線と脊椎動物の脊索が共通起源を持っている可能性を示唆する。しかしながら、節足動物の腹側正中線が外胚葉であるのに対して脊椎動物の脊索は中胚葉であることを考えると、その可能性に説得力を持たせるには胚葉の進化を理解することが必要不可欠である。
(4)At-delta とAt-notch は尾葉の外胚葉を特異化するのに必要である
昨年度までの解析によって、ショウジョウバエのtwist の相同遺伝子At-twist が将来の尾部領域から胚内部に入り込んでいる細胞で発現していることが分かっていた 4) 。本年度は、ショウジョウバエのDelta の相同遺伝子At-delta がAt-twist の発現に10時間ぐらい先行して将来の尾部領域の表面上皮細胞層で散在的な発現を示すことを明らかにした。そこで、At-deltaの役割を調べるためにpRNAiによってAt-deltaの機能抑制実験を行った。その結果、At-delta RNAi胚では尾部領域に普通には見られない大きな陥没ができ、その後の後体部の発生も大きく妨げられた。分子マーカーの発現解析から、At-delta RNAi胚で予定尾部領域から異常に陥入した細胞はすべてAt-twist を発現しており、逆に、本来尾葉の外胚葉で発現すべきショウジョウバエのcaudal の相同遺伝子At-caudal が発現しないことが分かった(図3)。このように異常になった尾葉では後方への伸長が見られず、後体部の体節も形成されなかった。さらに、その尾葉領域では細胞死も多数観察された。これらの結果は、At-delta がAt-caudal を発現する尾葉の外胚葉細胞を特異化するのに必要であることと、At-delta がAt-twist を発現する細胞の数を調節するのに関わっていることを示唆している。さらに、Deltaのレセプターをコードしていることが知られているNotch の相同遺伝子At-notch をクローニングし、その遺伝子の機能をpRNAiによって調べたところ、At-notch も予定尾葉領域でAt-delta と同じような役割を果たしていることが分かった(図3)。
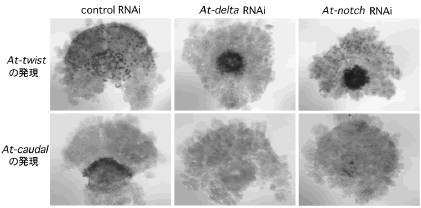 |
図3:At-delta とAt-notch のRNAiによる異常
予定尾葉領域においてAt-twist とAt-caudal の発現に異常が見られる
以上の結果を考え合わせると、Delta-Notch のシグナルによる側方抑制の仕組みが、At-twist を発現するようになる細胞とAt-caudal を発現するようになる細胞との間で働いていることが推測された。ショウジョウバエの胚発生では同じような仕組みが神経-表皮細胞間の運命決定に働いていることが知られているが5)、その仕組みはtwist を発現する初期中胚葉細胞やcaudal を発現する後端部の細胞の運命決定には全く関わっていない。この点においてショウジョウバエとオオヒメグモでは大きく異なっていた。また、オオヒメグモの予定尾葉領域はデフォルトではtwist を発現する中胚葉に運命づけられていることになるが、この中胚葉への誘導に必要な因子を探索することは今後の課題である。
おわりに
私たちは、複数の遺伝子の発現と機能の解析を通して、ショウジョウバエとオオヒメグモの発生システムの違いを分子レベルで具体的に明らかにしてきた。私たちの発見は、節足動物門内の進化においてかなり大胆な発生システムの変化があったことを示唆した。動物門を超えて動物の形態進化を議論するためには、私たちはもっとそれぞれの動物門の中の多様性を把握し、それぞれの動物門の祖先状態を慎重に吟味していく必要がある。