年度別活動報告
年度別活動報告書:2006年度
脊索動物と節足動物の共通祖先を理解する 2−2 カドヘリンの構造と機能の関係の解析
小田広樹(主任研究員) 秋山-小田康子(奨励研究員)
野田彰子(研究補助員) 春田知洋(大阪大学大学院生)
金山真紀(大阪大学大学院生)
はじめに
多細胞動物は細胞と細胞を連結するために特別な微細構造を持ち、それを巧みに操ることによって体の形を作り上げている。このような細胞間結合構造には何種類かのタイプが知られているが、アドヘレンスジャンクションは多細胞動物に最も普遍的に見られるジャンクションであり、形態形成に最も密接に関わっているジャンクションでもある。アドヘレンスジャンクションにおいて細胞と細胞を繋げている主要な分子はクラシックカドヘリン(以後、カドヘリンと呼ぶ)である。カドヘリンは隣接の細胞に発現する同種のカドヘリンを認識し結合する活性をもつ。私たちはこれまでの研究で、カドヘリンの細胞外領域のドメイン構成は左右相称動物の間で多様であり、その多様性はヒトデやクモで共通に見られる状態からの独立的な短縮化で説明できることを見いだした8)。短縮化されたあるひとつの状態(昆虫型)は、ハエ、コオロギ、トビムシ、アルテミアで見られ、それとは別の状態(脊椎型)が脊椎動物とホヤで、そして、さらに別の状態(頭索型)がナメクジウオで見られる。それぞれの状態は進化的に安定であるが、動物の初期進化のある時点では大きな変化があったと考えられる。私たちは、そのようなカドヘリンの構造変化がカドヘリンの機能にどのような変化をもたらしたのかを実験データに基づいて検証したいと考えている。そこでまず、ジョウジョウバエのカドヘリン(DEカドヘリン)を用いて、昆虫型カドヘリンのドメイン構成がなぜ進化的に安定に保持されているのか、言い換えると、昆虫型カドヘリンの構成要素それぞれが果たす必要不可欠な役割は何かを調べた。とりわけ、脊椎型カドヘリンのドメイン構成との違いに重点を置いて解析を行った。
DEカドヘリンは、細胞外領域に7個のカドヘリンドメイン(EC1-7)と、無脊索ドメイン(NC)、システインリッチEGF様ドメイン(CE)、ラミニングロビュラードメイン(LG)で構成される原始クラシックカドヘリンドメイン(PCCD)複合体を持つ(図4)。昨年度までの解析では、様々なドメインを欠失したDEカドヘリン分子(すべてGFPがタグとして付加されている)を作製し、それらの分子が培養細胞でどれだけの接着活性を示すかを調べた。その結果、EC7とPCCD複合体をともに欠失した分子(DE-EC6と呼ぶ、図4)でも正常分子と同程度の接着活性を示すことが明らかになっていた。そこで本年度は、このDE-EC6が個体内の発生現象においてどれだけの機能を発揮できるかを調べた。
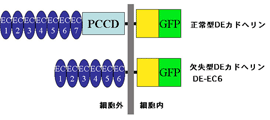 |
図4. 正常型DEカドヘリンと作製した欠失型DEカドヘリンの模式図
結果と考察
DE-EC6は正常分子と同じようにアドヘレンスジャンクションに局在することができる
UASプロモーターまたはユビキチンプロモーターの制御下でDE-EC6を発現するトランスジェニックショウジョウバエ系統を作出し、胚にDE-EC6を発現させたところ、DE-EC6は正常分子と同じように上皮細胞のアドヘレンスジャンクションに局在した。DEカドヘリンの突然変異体shotgun(shg)のバックグラウンドにDE-EC6を発現させてもアドヘレンスジャンクションに局在した。過去に記述されたPCCD複合体に変異をもつDEカドヘリン分子とは異なって、DE-EC6は生体内の上皮細胞で正常にふるまうことができることがわかった。
DE-EC6は正常分子に近いレベルのトラキア(気管)融合活性をもつ
shgヌル変異体では、袋状に陥入したトラキア原基が隣接の体節間で融合しない9)。このshg変異体において、トラキア細胞だけにDE-EC6を発現させてトラキアの融合不全を救済できるかを調べたところ、90%以上の融合ポイント(1個体当たり20ポイント)において完全な救済が認められた(図5)。正常型DEカドヘリン分子では100%に近い融合ポイントで異常が救済されたが、細胞質領域を持たないDEカドヘリン分子やEC2-3に欠失を持つ分子ではそのような救済は全く見られなかった(図5)。これらの結果は、DE-EC6が正常分子に近いレベルのトラキア融合活性をもつことを示した。EC7とPCCD複合体がなくてもDEカドヘリンは生体内である程度の機能を発揮しうることがわかった。
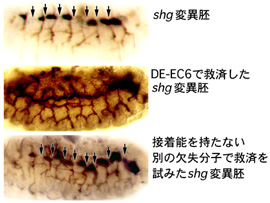 |
図5.shg変異体を用いたトラキア融合不全救済実験. 矢印はトラキアが体節間で融合できていないポイントを指している.
おわりに
EC7とPCCD複合体を欠いたDE-EC6は、培養細胞による集合実験においても、トラキア融合不全救済実験においても、正常分子と同等レベルまたはそれに近いレベルの活性を示した。DE-EC6はDEカドヘリンのすべての機能について正常型DEカドヘリンの肩代わりをできるのであろうか? それとも、DE-EC6では肩代わりできない機能が存在するのであろうか? 現在、ユビキチンプロモーターの制御下でDE-EC6を発現させてshg変異体の救済実験を行っている。もし正常型カドヘリンでは遂行できるが、DE-EC6では遂行できないような機能を見つけることができれば、無脊索動物に特異的な領域であるPCCD複合体の役割を理解するための助けになるであろう。これらの解析を通して、無脊索動物と脊索動物の上皮細胞の違いを理解することができるのではないかと期待している。
参考文献
1) Carroll, S. B., Grenier, J. K., and Weatherbee, S. D. (2001) From DNA to diversity. Blackwell Science.
2) Oda, H., Wada, H., Tagawa, K., Akiyama-Oda, Y., Satoh, N., Humphreys, T., Zhang, S., and Tsukita, S. (2002) A novel amphioxus cadherin that localizes to epithelial adherens junctions has an unusual domain organization with implications for chordate phylogeny. Evolution & Development 4, 426-434.
3) Delsuc, F., Brinkmann, H., Chourrout, D. and Philippe, H. (2006). Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 439,965-968.
4) Oda, H., Nishimuta, O., Hirao, Y., Tarui, H., Agata, K. and Akiyama-Oda, Y. (2007). Progressive activation of Delta-Notch signaling from around the blastopore is required to set up a functional caudal lobe in the spider Achaearanea tepidariorum. submitted.
5) Hooper, J. E. and Scott, M. P. (2005). Communicating with Hedgehogs. Nat. Rev. Mol. Cell Biol. 6,306-317.
6) Akiyama-Oda, Y. and Oda, H. (2003) Early patterning of the spider embryo: A cluster of mesenchymal cells at the cumulus produces Dpp signals received by germ disc epithelial cells. Development 130, 1735-1747.
7) Akiyama-Oda, Y. and Oda, H. (2006) Axis specification of the spider embryo: dpp is required for radial-to-axial symmetry transformation and sog for ventral patterning. Development 133, 2347-2357.
8) Oda, H., Tagawa, K. and Akiyama-Oda, Y. (2005) Diversification of epithelial adherens junctions with independent reductive changes in cadherin form: identification of potential molecular synapomorphies among bilaterians. Evol. Dev. 7, 376-389.
9) Uemura, T., Oda, H., Kraut, R., Hayashi, S., Kotaoka, Y. and Takeichi, M. (1996). Zygotic Drosophila E-cadherin expression is required for processes of dynamic epithelial cell rearrangement in the Drosophila embryo. Genes Dev. 10,659-671.