年度別活動報告
年度別活動報告書:2003年度
脊索動物と節足動物の共通祖先を理解する 4-1 細胞間結合構造に基づいた動物の系統関係の推定
小田広樹(研究員)
秋山-小田康子(派遣研究員)
山崎一憲(奨励研究員)
入江雅美(派遣実験補助員)
田中千咲(筑波大学生)
はじめに
一般的に広く行われている、DNAの塩基置換に基づいた系統解析は、動物の門や綱のレベルの(動物進化の初期で起こった)系統分岐パターンを再構成するには必ずしも適していない1), 2)。なぜならDNAの塩基置換には動物間における置換速度のばらつきや分子内の置換速度のばらつきなどが存在し、これらの影響によって分岐点が深くなればなるほど推定の確率論的確からしさが低くなるからである。
そこで近年は、このような従来の系統解析の方法に代わって、長い進化の歴史の中で稀にしか起こらなかった遺伝的な変化を見出すことによって、“深い”系統分岐をより確からしく推定しようとする試みがなされている3)。言い換えれば、現存動物のゲノム中に見られる相同形質において、ある動物群に共有されている派生的状態(Synapomorphy)を見つけ出すことによって、それを共通子孫を知るためのマーカーに使おうとする試みである。しかしながら、この方法においてもホモプラシー(二次的に類似性が生じること)や原始的状態と派生的状態を見分ける際の客観性などに問題があり、多細胞動物の深い系統関係を解く有効な形質状態を見出すことは一般的には難しい。
本研究は、アドヘレンスジャンクション(adherens junction, AJ)と呼ばれる細胞間結合構造が左右相称動物の間で多様な構造的状態をとっていることに基づいて、その形質状態が多細胞動物の深い系統関係を知るために有効な情報となりうるかを検討した。本年度は昨年度までのデータにさらに新しいデータを加えて解析し、左右相称動物におけるAJの原始的状態と派生的状態を判定した。そして、派生的状態の分布パターンから脊椎動物、尾索類の近縁性及び昆虫類、トビムシ類、鰓脚類の近縁性が支持された。
結果と考察
昨年度までに本研究で決定した左右相称動物のカドヘリン及びすでに公的データベースに記載されているカドヘリンの構造を7つのタイプ (A1、A2、E1、E2、H1、C、V) に分類した(図1)。そして、初期胚の上皮細胞でAJの形成を担っているカドヘリンの構造タイプによって、それぞれの動物種のAJの構造状態を定義づけた。これまでに構造決定が完了しているが、上皮細胞のAJ形成への関わりが不明なカドヘリンに関しては、特異抗体を作製してそのカドヘリン蛋白質の発現を調べた(図2)。それにより、各動物種のAJの構造状態を特定し、その結果を図3の中にまとめた。コオロギ(バッタ類)、トビムシ、アルテミア(鰓脚類)のAJはショウジョウバエと同じ状態A1であり(図2C, D)、フナムシ(等脚類)、エビ(十脚類)、クモ(鋏角類)とは状態が異なっていた。後者のグループでは、抗体染色による解析からA2タイプのカドヘリンが上皮のAJ形成を担っていることがわかり(図2H, J, K)、節足動物においては主に状態A1と状態A2の、2つの構造状態があることが示された。ただし、前者のグループにもA2タイプのカドヘリンは存在している。しかし、そのカドヘリンはショウジョウバエで知られているように主に非上皮組織に存在していた(図2E, M, N, O)。節足動物におけるこれらの一連のデータは、AJの構造状態が進化の過程で稀にしか変化しないものであることを支持し、AJの解析が左右相称動物の深い分岐を解くために重要な情報を与えうることを示唆した。
|
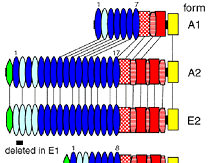
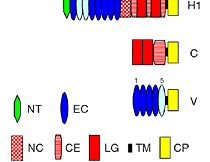
図1:左右相称動物に見られるAJの構造的状態.
AJの構造的状態は上皮細胞のAJ形成を担うカドヘリンの構造タイプによって定義する。
|
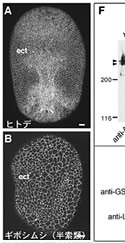

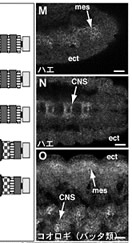
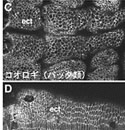
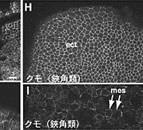

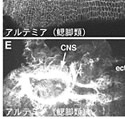
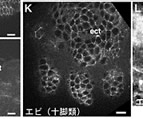
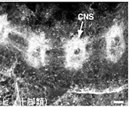
図2:様々な動物の外胚葉性上皮におけるカドヘリンのAJへの局在
(A-E) 特異抗体による染色。(F) 抗At-cadherin抗体、抗Le-cadherin抗体によるウエスタン解析。(G) 抗Le-cadherin抗体のA2型cadherinに対する交差反応. (H-O) 抗Le-cadherin抗体による染色.
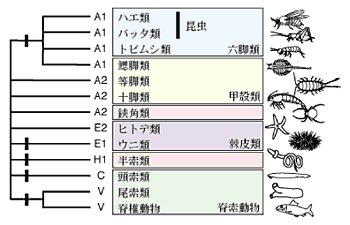 |
図3:各動物のAJの構造状態と推測される動物の系統関係.
黒いボックスは原始的状態から派生的状態への変化を示している。異なる派生的状態が段階的に生じた可能性も否定できないが、この図はすべての派生的状態がA2またはE2様の状態から直接生じた可能性が高いという私たちの仮説的考えに基づいている。
そこで次に、状態A1と状態A2のどちらが原始的で、派生的なのかを調べるために、A1タイプのカドヘリンとA2タイプのカドヘリンの各ドメインのアミノ酸配列をBLAST配列アライメントツール4)を活用して比較した。その結果、A1タイプのカドヘリンのEC1からEC6, EC7はA2タイプのカドヘリンのEC8からEC13、EC17に対応することが示唆され(図1のドメイン間の線)、A2タイプからドメインの欠失によってA1タイプが生じた可能性が高いことがわかった。そしてさらに、A2タイプのカドヘリンは棘皮動物ヒトデのE2タイプのカドヘリンとの間に、アミノ酸配列によって特徴づけられる個々のドメインの構成及び配置に関して保存性が認められ(図1)、節足動物の状態A2と棘皮動物の状態E2の類似が二次的なものではなく、左右相称動物の共通祖先の状態をそのまま引き継いできた結果であることが示唆された。それに対し、昆虫、トビムシ、アルテミアで見られる状態A1は節足動物の進化過程で生じた派生的な状態であると考えられた。この考えは、ゲノム情報が蓄積しているにも関わらずA1タイプのカドヘリンが節足動物以外の動物で見つかっていないという現状とも辻褄が合う。
(2)脊椎動物、尾索類の近縁性と昆虫類、トビムシ類、鰓脚類の近縁性
状態A1とは異なるAJの派生的構造状態は、脊椎動物、尾索類、頭索類、半索類、ウニ類で見られる。系統関係を解く上では、原始的状態から派生的状態への変化がどのようなパターンで起こったかが重要な鍵となる。私たちは、起こりにくいはずの状態変化がある系譜で複数回起こって派生的状態が段階的に生じたという可能性よりも、原始的状態から派生的状態への直接変化がいくつかの異なる系譜で独立に起こった可能性の方が高いのではないかと仮説的に考えた。この仮説に基づくならば、図3で示したように派生的状態の分布パターンによって脊椎動物、尾索類の近縁性と昆虫類、トビムシ類、鰓脚類の近縁性が支持され、頭索類や等脚類、十脚類はそれらの2つの動物グループの外に位置づけられることになる。このような系統関係は、塩基置換に基づいた系統解析の結果とは必ずしも一致しない5),6),7)。どちらが“真”の系統関係を示しているのか、またどちらも正しくないのか、AJとは独立の形質を調べることによって最も辻褄の合う系統関係を丹念に検討していかなければならない。
|
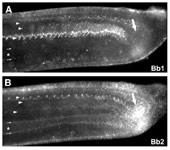
(3)ナメクジウオのカドヘリンの解析
昨年度までに私たちは2つのナメクジウオのカドヘリンをクローニングし、その遺伝子の塩基配列を決定した。それらのカドヘリンはBb1カドヘリン(Bb1C)、Bb2カドヘリン(Bb2C)と命名された。Bb1CはOda et al., 2002に報告したものである8)。図1に示したように、Bb1CとBb2Cはお互いによく似た構造をとっているが、それらの細胞外領域の構造は他の動物のカドヘリンと比べると極端に異なっていた。つまり、ナメクジウオのカドヘリンはカドヘリンの接着活性に必須だと考えられているECドメインが全くなかった。本年度は、このようにユニークな構造をしたBb1CとBb2Cが脊椎動物やショウジョウバエで知られているカドヘリンと同じような様式で細胞接着及び胚の形態形成に関与しているかどうかを検討した。ショウジョウバエ培養細胞を用いた集合実験では、Bb1CとBb2Cはいずれもカルシウムイオン非依存性の接着分子として働き、それぞれのカドヘリンに接着特異性があることが明らかになった。そして、Bb1CとBb2Cに対する特異抗体を用いたナメクジウオ胚の二重免疫染色では、それらのカドヘリンが異なる発現パターンを示し(図4)、多層細胞からなる構造の発生に関与している可能性が考えられた。このように構造がユニークであってもBb1CとBb2Cは、脊椎動物やショウジョウバエで知られているカドヘリンと同じように胚の形態形成に重要な貢献をしていることが推察された。この研究は本年度論文にまとめた9)。
おわりに
私たちは、多細胞動物にとって欠かせないアドヘレンスジャンクションという細胞間結合構造を調べることによって動物の系統関係を再構成するための手がかりを得た。しかしながら、分岐が深かろうが浅かろうが系統関係を結論づけることは極めて難しい。そこで本研究が科学的見地から重視すべき点は、ある動物群に保存された特異的な状態を見出し、その機能的な意味を理解することにあると考えている。今後も、細胞間結合を制御する細胞システムの解析を通じて脊椎動物と昆虫に向かった動物の進化を追究していきたい。