年度別活動報告
年度別活動報告書:2003年度
3.昆虫と植物との共生関係、共進化および種分化 3-2. オオバギ - シリアゲアリ共生系における種特異性の創出・維持、認識機構
蘇 智慧(研究員、代表者)
村瀬 香(奨励研究員)
神田嗣子(研究補助員)
中村桂子(館長)
はじめに
世界中の熱帯域では、アリ類に被食防衛をゆだねている植物が数多く報告されている。その中には、特定のアリ類に体の一部を巣場所として提供して、そのアリ類と共生している植物が知られている。このような植物をアリ植物と呼ぶ。アリ植物と共生する以外には生息できないほどアリ植物に特殊化したアリ類を、植物アリ(共生アリ)と呼ぶ。両者は、お互いなしでは生存できないほど相手に依存している絶対的な相利共生関係にあることが知られている。
東南アジア熱帯域を中心に分布するオオバギ属(Macaranga: Euphorbiaceae,トウダイグサ科)は、こうした絶対共生型のアリ植物種を数多く含んでいる。オオバギ属のアリ植物は共生アリに巣場所を提供するだけでなく、餌(栄養体)も提供している。一方共生アリは、栄養体を主な餌資源として利用し、植物を植食者から防衛し、植物上のみで生活をしている。オオバギ属のアリ植物の共生相手となるアリ種の大半は、シリアゲアリ属(Crematogaster: Myrmicinae)に属している。オオバギ-シリアゲアリ系では、1種のアリ植物は、その種のみあるいはその種の近縁種を含む少数の種群のみに特殊化した1-2種のシリアゲアリ種と共生関係をもつことが分かっており、高い種特異性が創出・維持されている。
種特異性の共生系が創出・維持される機構があるならば、崩壊する機構もあるはずである。種分化や多様性創出機構を考えるには、種特異性が崩壊する機構も調査する必要がある。そこで、原生林から二次林まで、広範囲に生息しているアリ植物種に注目した。原生林と、人為的撹乱などで生じた二次林とでは、光条件などの環境条件が全く異なる。生息環境条件が異なっていても、種特異性は維持されているのかを調査した。
結果と考察
オオバギ属は、南太平洋諸島から西アフリカまで約300種が分布している。マレーシア(ボルネオ島およびマレー半島)では、52種以上のオオバギ種が報告されており、そのうちの25種以上がアリ植物である。本研究におけるすべての野外調査、サンプリングは、東南アジア熱帯域に属する熱帯低地林(マレーシア国、ボルネオ島、サラワク州、ランビルヒルズ国立公園)で行った。調査の結果、形態的な特徴は原生林の共生アリ種と同じであるが、その行動や生活史が異なるアリが二次林から見つかった。そこで、これらのアリからDNAを抽出し、ミトコンドリアCO1遺伝子を用い系統解析を行ったところ、二次林から採集された個体は2つのグループにわかれた(図4)。そのうちの1つは原生林のものと極めて近縁だが、もう1つのグループはかなり異なっていた。このことから、植物の生息環境の変異性が新たな種間関係を創出する機構となりうることが示唆された。しかし、サンプルの調査地や分析したアリの数が極めて限られているので、微小環境の状態、植物の状態、アリの攻撃性、女王の有無といった生態学的調査を含め、更なるDNA解析も必要である。
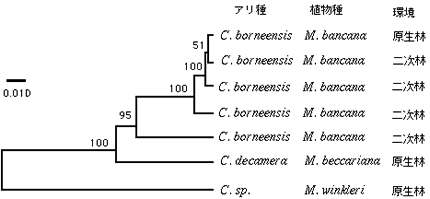 |
図4 二次林の共生アリの系統樹。
おわりに
イチジク属とイチジクコバチの関係は寄生コバチを含めた三者の関係が絡み合った複雑な系である。イチジク属と送粉コバチの共生関係の進化を探る上ではこれら三者の生活史がどのように関連しているかを知る必要があるが、まだほとんど分かっていないのが現状である。寄生コバチは宿主の転換によって種分化を起こしているようだが、その転換の機構と条件について更なる研究が必要である。送粉コバチはイチジクとの「1種対1種」の関係がかなり安定している。実際に共進化をしていたかどうかを語るためには、更に材料を増やす必要がある。また、日本産イチジクとイチジクコバチの起源や列島への進入解明にも周辺地域のサンプルを解析しなければならない。今後も DNA 解析と生態調査などを通してイチジクとイチジクコバチの共生・共進化の謎を解いていきたい。
アリ植物と共生アリとの共生関係については、今後、様々な生態学的なデーターとリンクしたサンプルを地道に集め、分子系統解析を行い、シリアゲアリ属の多様化の機構やオオバギ植物への特殊化の進化過程を明らかにしていきたい。
引用文献
1) Rasplus J-Y, Kerdelhue C, Clainche I L, Mondor G (1998) Molecular phylogeny of fig wasps Agaonidae are not monophyletic. C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 321: 517-527.
2) Weiblen, G. D. (1999) Phylogeny and ecology of dioecious fig pollination. Ph. D. Thesis, Harvard Univ., Cambridge, Massachusetts.
3) Yokoyama J (1995) Insect-plant coevolution and speciation. In R. Arai, M. Kato, and Y. Doi [eds.], Biodiversity and evolution, 115-130, National Science Museum Foundation, Tokyo, Japan.
4) Yokoyama J, Iwatsuki K (1998) A faunal survey of fig-wasps (Chalcidoidea: Hymenoptera) distributed in Japan and their associations with figs (Ficus: Moraceae). Entomol. Sci. 1: 37-46.
5) 蘇智慧, 東浩司 (2002) 日本産イチジクコバチの分子系統と進化. 昆虫と自然 37: 8-12.
6) 東浩司, 蘇智慧, 中村桂子 (2003) イチジク属とイチジクコバチの共進化. 数理科学 No. 479: 78-83