年度別活動報告
年度別活動報告書:2004年度
3.昆虫と植物との共生関係、共進化および種分化 3-1. イチジクとイチジクコバチの共進化と種分化を探る
蘇 智慧(研究員、代表者)
佐々木剛(奨励研究員)
神田嗣子(研究補助員)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士或いは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物はそれぞれ陸上で最も多様化した生物群で、その多様化は植物と昆虫とが互いに相互適応的関係を築くことによって促されてきた。したがって、植物と昆虫との相互関係(競争、共生、寄生など)を解明することは、生物の多様性を理解する上で最も重要なカギの一つである。我々は現在植物と昆虫の共生関係の中で最も代表的な系といわれるイチジク属とイチジクコバチの共生関係についてDNA系統解析や野外観察調査などにより研究を行っている。
イチジク属Ficusは、クワ科(Moraceae)に属し、4亜属(イチジク亜属Ficus, アコウ亜属Urostiguma, Pharmacosycea, Sycomorus)に分けられ、熱帯を中心に約750種が世界中に分布している。日本には南西諸島を中心にアコウ亜属3種、イチジク亜属13種が分布している。一方、イチジクコバチの仲間は分類学的に膜翅目、コバチ上科Chalcidoideaに属し、イチジクコバチ科Agaonidaeとして分類されている。イチジクコバチ科はさらに複数の亜科に分けられ、その内の1つは送粉コバチ亜科Agaoninae (pollinating fig-wasps)で、他の亜科はすべて寄生コバチによって構成されている1)。送粉コバチとイチジクとの間には、子孫を残すための共通利益のもとで、「1種対1種」という絶対的な相利共生関係が結ばれていると言われている。しかし、寄生コバチの場合は、複数種の寄生コバチが同種のイチジクに寄生することができる。イチジクとイチジクコバチとの関係は古くから共進化のモデル系として多くの研究をなされてきた。特に近年アフリカや南米の材料を中心に分子系統解析の研究も盛んに行われるようになった2)。日本産イチジク属とイチジクコバチについても研究が行われている3, 4)。
我々はこれまでメキシコ産、日本産と中国海南島産のイチジクとイチジクコバチの分子系統解析を行ってきた。メキシコ産の材料を分析したところ、Urostigma亜属の送粉コバチはおよそ2100万年前に一斉放散分化したことが示唆され、また、同じ種のイチジクから採集した送粉コバチが系統樹上1つの枝にまとまらず、異なる系統に分かれるという興味深い結果が得られた5)。日本産の材料を調べた結果、イチジクと送粉コバチの「1種対1種」の関係はほぼ厳密に維持されており、系統解析の結果は部分的に共進化を支持した。また、寄生コバチの多様化・種分化においても多くの知見が得られた。ところが、期待していた琉球列島間のイチジクコバチの地域的変異は全く見られなかった6, 7)。今年度は中国海南島産のイチジクとイチジクコバチの分子系統解析を行うと同時に、核rRNA internal transcribed spacer (ITS) という分子マーカーを駆使してイチジク属植物の系統解析を行い、より詳しい系統関係が解明され、イチジク植物と送粉コバチとの共進化がより明瞭となった。また、中国産アコウとガジュマルの送粉コバチの解析によって、これらのイチジク植物に複数の系統(種?)の送粉コバチが発見された。
結果と考察
1) ITSによる日本産イチジク植物の系統解析
これまで葉緑体DNAを用いて日本産イチジク属植物の系統解析を行った結果、部分的に送粉コバチの系統関係と一致し、両者が共進化したことが示唆されたが、植物の系統関係においてまだ不明瞭な点が残っていた。例えば、日本産イチジク各種が葉緑体DNAの系統樹上、5つのグループにまとまるが、その5つのグループの分岐順序がはっきりしない。また、グループ内においても、オオバイヌビワF. septica、アカメイヌビワF. beguentensis、ギランイヌビワF. variegata 3種の系統関係は明らかでない7)。そこで、今回は核 ITS領域を用いて、それらの系統関係を再検討した。その結果を図1に示す。葉緑体DNAの系統樹でははっきりとしなかったオオバイヌビワ、アカメイヌビワ、ギランイヌビワ3種の系統関係は、核ITSの解析によりはじめて明らかになった。オオバイヌビワとアカメイヌビワが近縁で、ギランイヌビワはそれらの外群である。また、新しい結果として注目に値するのは、オオイタビF. pumila/ヒメイタビF. thunbergii/イタビカズラF. nipponicaとイヌビワF. erecta/トキワイヌビワF. boninsimaeが同一クラスターを形成し、共通祖先を示した点である。これらの結果は送粉コバチの系統関係とより対応することになり、イチジクとイチジクコバチの共進化をさらに強く支持した。葉緑体と核ITS領域を併せて作成した系統樹と送粉コバチの対応関係は図2に示す。しかし、残念ながら、大きいグループ間の関係は解明されなかった。これは今後更なる研究が必要である。
2) アコウやガジュマルに複数の送粉コバチの系統
すでに述べたように、イチジク属植物とイチジク送粉コバチの間では、「1種対1種」という極めて厳密な共生関係があると言われている。この関係をDNA系統解析によって確認するには、各種の分布域を広くカバーするようなサンプリングと、緻密な解析を行うことが必要である。日本産イチジク属植物の多くの種においては、様々な遺伝子を用いて、このような系統解析を行った結果、1種のイチジク植物の花嚢から複数の異なる系統の送粉コバチを得ることはできず、「1種対1種」の関係は確認された。しかし、アコウF. superbaとガジュマルF. microcarpaといった雌雄別株の種に関しては、複数の送粉コバチの系統が認められた(図3)。これらの系統間の遺伝的違いは、種間変異を超えるほど全く異なるものであることが判明した。また、日本産アコウの花嚢から採集した送粉コバチは、そのうちの1系統は中国海南島産アコウの送粉コバチに極めて近縁であることも分かった。一方、これらの送粉コバチの異なる系統の宿主植物のDNA解析を行ったところ、遺伝的な変異は全く見られなかった。このことは2種のイチジク送粉コバチが同種のイチジクに送粉していることを強く示唆している。これら2種(?)の送粉コバチは、同種の宿主に対して全く同じ認識機構を持っているかどうかが興味深い。もしかすると宿主側には送粉コバチに認識される化学物質などのシグナルが微妙に異なっているかもしれない。それにしても、宿主側には全く遺伝的変異が見られないことを考えると、両者の種(系統)分化が同調して起きていたとは考えられない。これら2種(系統)の送粉コバチがどうやって分岐したのか?興味深い課題である。
また、中国産と日本産の送粉コバチとの間の地域差が見られたことから、日本産イチジク送粉コバチの種内地域変異が全くないことは遺伝的交流が広範囲にわたって行われているためであることが示唆された。
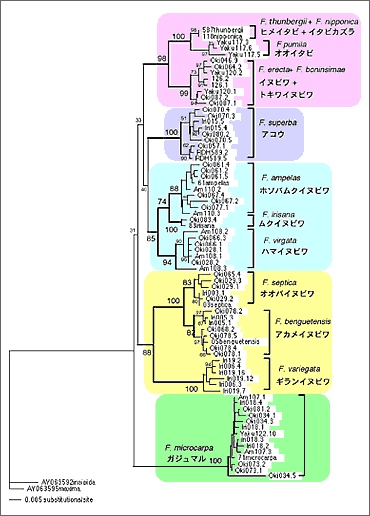 |
図1. 核ITS領域による日本産イチジク属植物の系統樹。枝の分岐点にある数字は系統樹の信頼度を示すBootstrap値である。植物の種名(学名と和名)は系統樹の右側に示してある。13種のイチジク植物が種ごとにclusterを形成し、更に5つのグループにまとまっている。グループ間の系統関係ははっきりしないが、グループ内の種間関係(種の分岐順序)は明らかになっている。
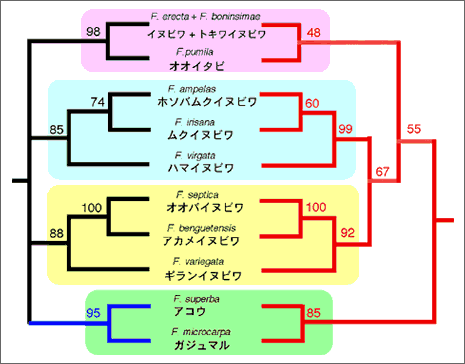 |
図2. 葉緑体DNAと核ITS領域による日本産イチジク属植物の系統樹(左)とミトコンドリアCOI遺伝子によるそれらの送粉コバチの系統樹(右)の対応関係。枝の上の数字は系統樹の信頼度を表すBootstrap値である。両系統樹の間の和名と学名は植物の名前である。植物の方は、4つのグループにまとまっており、グループ内の種間関係も綺麗に解明されている。これに対して、送粉コバチの方はほぼ同様な系統関係を示しており、両者間は同調して種分化していることが示唆されている。
 |
図3. 核28SrDNAによるアコウとガジュマルの送粉コバチの複数の系統を表す系統樹。系統樹の各枝の上或いは下にある数字はBootstrap値で、枝先の学名はコバチの名前である。各コバチの宿主植物の名前は系統樹の右側に示してある。系統樹からアコウとガジュマルの送粉コバチはそれぞれ大きく2つの系統に分かれていることが分かる。これら2つの系統の分岐はオオバイヌビワコバチとアカメイヌビワコバチの種分化(系統樹の一番下の2種)よりも古い。四角で囲んでいるアコウとガジュマルの送粉コバチの系統は恐らくそれぞれ別種(新種?)と考えられる。