年度別活動報告
年度別活動報告書:2005年度
3-2. 昆虫類(六脚類)の起源、系統と進化に関する研究
蘇 智慧(研究員、代表者)
佐々木剛(奨励研究員)
石渡啓介(大学院生)
神田嗣子(研究補助員)
はじめに
陸上昆虫類(六脚類 Hexapoda)は、無翅昆虫類と有翅昆虫類とに大きく分けられる。無翅昆虫類はさらに4目(カマアシムシ目、トビムシ目、コムシ目、シミ目)、有翅昆虫類は26目に分類されている。従来、六脚類は他の節足動物のうち多足類(Myriapoda)と近縁であり、六脚類内部では無翅昆虫類から有翅昆虫類のグループが分岐し翅を獲得したと考えられてきたが、最近の分子系統解析により、六脚類は多足類ではなく甲殻類(Crustaceae)と近縁であることが明らかになった。さらに六脚類の単系統性にも疑問がもたれ、Nardi ら 7) によるミトコンドリア遺伝子を用いた研究では、シミ目を含む有翅昆虫類は甲殻類と近縁であり、無翅昆虫類のトビムシ目はより古い時代に分岐したという可能性も示唆されている。しかし、この結果は解析方法によって変わるなど有意なものではなく、トビムシ目以外の無翅昆虫類も含まれていない。Regier ら8)は、核にコードされたタンパク遺伝子RNA polymerase II largest subunit (RPB1), elongation factor 1α (EF-1α), elongation factor 2 (EF-2)を用いた解析により六脚類の単系統性を示したが、EF-1α 遺伝子を用いるのには問題がある(後述)。また、彼らのデータには無翅昆虫類のカマアシムシ目が含まれていないため、六脚類の単系統性には疑問が残されているのが現状である。
そこで私たちは多数の核ゲノム上のタンパクをコードしている遺伝子を比較して無翅昆虫類の正しい系統的位置を解明し、昆虫類の起源、系統と進化を明らかにし、六脚類の共通祖先が一度だけ陸上へ進出したのか、それとも複数の系統の祖先が独立に複数回上陸したのかという問いに答えることを目的として、昨年度後半より本研究を行っている。
結果と考察
1) 系統解析に適した遺伝子の探索
これまでに、18S ribosomal RNA (18S rRNA), Elongation Factor 1 alpha (EF-1α) および 2 (EF-2), RNA polymerase II largest subunit (RPB1), ミトコンドリアゲノム遺伝子などが系統解析に用いられてきた。18S rRNA 遺伝子は、データは豊富にあるが、系統間の進化速度の違いが大きく、系統解析には向かない。ミトコンドリア遺伝子も同様である。そこで、核にコードされたタンパク遺伝子を検討した。EF-1α 遺伝子は、六脚類において遺伝子重複が観察され、パラロガスな比較をしてしまう危険性があるため六脚類の系統関係を推定するのに用いることはできない。これまでに用いられてきたRNA polymerase II largest subunit (RPB1) に加えて、新たに RNA polymerase II second largest subunit (RPB2) DNA polymerase delta catalytic subunit (DPD1) 遺伝子が、系統解析に適していることがわかった。それは、これらの遺伝子に、
(1)節足動物の系統内部で遺伝子重複がない。
(2)適度な保存性を有しており、縮退プライマーの設計が可能である。
(3)PCRの結果が良好である。
といった性質があるためである。
2) 3つの核タンパクコード遺伝子(RPB1, RPB2, DPD1)による系統解析
専門家やアマチュアの方々の協力を得て無翅昆虫類、有翅昆虫類、そして甲殻類や他の節足動物の材料を入手した。RNAを抽出して逆転写を行い、cDNA を鋳型にして縮退プライマーを用いたPCR反応を行い、塩基配列決定を行っている。また、RACE法により可能な限り各遺伝子のアライメント可能な領域全長配列を決定している。さらに、最尤法による分子系統解析を行うため、7台のコンピュータによる並列計算環境を構築した。
これまでに得られたデータから最尤法に基づく分子系統解析を行った結果、無翅昆虫類のなかでもシミ目は有翅昆虫類と近縁となり、外顎類(有翅昆虫類+シミ目)をひとつの分類群とする考え方が強く支持された。シミ目に属するイシノミとシミは単系統とはならず、有翅昆虫類とシミが近縁でありイシノミはより古くに分岐したことが支持された(図5)。内顎類の各グループ(カマアシムシ目、トビムシ目、コムシ目)の分岐順序ははっきりしなかった。有翅昆虫類内部では、トンボとカゲロウの旧翅類が他の新翅類より古くに分岐したことが支持された。さらに、進化速度が極端に大きい系統をのぞいた解析により、強い支持は得られなかったものの、鰓脚類は六脚類全ての姉妹群となり、六脚類は単系統である可能性が高いことが示唆された(図6)。
3) 「1種対1種」関係の厳密性と例外発見
イチジク属植物と送粉コバチの「1種対1種」関係について、これまでの解析結果から、相当厳密に維持していることがはっきりした。同種のイチジク属植物から採集された送粉コバチは、系統樹上乱れることなく、同じ配列を示すか、僅かな違いがあるにしても綺麗にまとまっていた。しかし、興味深い例外が見つかっている。ギランイヌビワの花嚢から採集した送粉コバチはアカメイヌビワコバチと全く同じ配列を示し、またアカメイヌビワとイヌビワから採集した送粉コバチが、逆にギランイヌビワコバチと同配列だったのである。これは、アカメイヌビワコバチがギランイヌビワの花嚢に進入でき、逆にギランイヌビワコバチがアカメイヌビワとイヌビワの花嚢に進入できることを意味する。言い換えれば、送粉コバチはある環境下(例えば、本来の宿主種がまわりにいない)において、別種の花嚢にも入ることが可能である柔軟性を持っていることが示唆された。但し、これらのコバチはすべて親コバチであるため、産卵できたかどうか、子どもコバチができたかどうかは確認されていない。
4) 小笠原諸島の固有種と移入種の起源
前にも述べたように、小笠原諸島には固有種3種(トキワイヌビワ、オオトキワイヌビワとオオヤマイチジク)と移入種1種(ガジュマル)が分布している。オオヤマイチジクを除いた他の3種の材料は採集することができた。解析の結果によると、トキワイヌビワとオオトキワイヌビワは全く区別がつかないほど近縁なものであり、それらの送粉コバチも同様差異はなかった。また、これら2種は、植物も送粉コバチも、それぞれ本州と南西諸島に広く分布しているイヌビワおよびその送粉コバチと近縁であることが分かった。オオヤマイチジクについては、植物乾燥標本を用いた一部の解析結果から、トキワイヌビワ、オオトキワイヌビワとイヌビワに極めて近縁であることが示唆された。これはYokoyama 1) の結果とも一致する。以上のことから、小笠原諸島の固有種3種は、南西諸島(或いは本州)から進入したイヌビワが島内で種分化したものであると考えられる。但し、トキワイヌビワとオオトキワイヌビワは本当に2種にすべきか、再検討する必要がある。
移入種のガジュマルは明治初期に防風林として移入・栽培されたものである。長い間、結実していなかったが、近年実が付くようになったと言われている。換言すれば、ガジュマルは近年になってようやく受粉できるようになったのである。その送粉コバチの正体は興味深い。 トキワイヌビワなど3固有種の送粉コバチがガジュマルにも送粉するようになったのか(宿主の転換)、或いは本来のガジュマルコバチが近年移入されたのかを調べたところ、小笠原のガジュマルコバチは南西諸島のものと全く同じであることが分かった。つまり、もし起きていれば面白いと思った宿主転換ではなく、人為的にガジュマルコバチが近年になって移入されたのだろう。この事実からも、イチジクとイチジクコバチの「1種対1種」関係は非常に堅いものと思われる。
おわりに
イチジク属とイチジクコバチの関係は寄生コバチを含めた三者の関係が絡み合った複雑な系である。イチジク属と送粉コバチの共生関係の進化を探る上ではこれら三者の生活史がどのように関連しているかを知る必要があるが、まだほとんど分かっていないのが現状である。今後は、送粉コバチの上位分類群の系統関係解明、非送粉コバチの詳しい系統関係、中国産イチジク属植物とイチジクコバチの系統関係および日本産のものとの繋がりなどを中心に解析を行い、イチジクコバチによる宿主転換の可能性や転換機構および種分化に与える影響などに関する理解を深めたい。
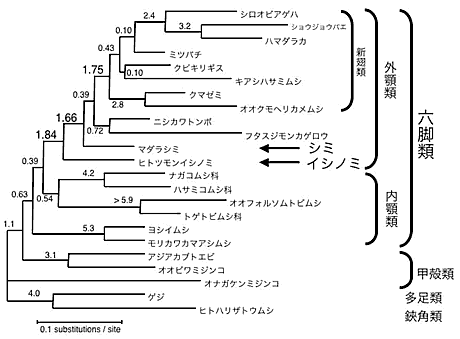 |
図5. DPD1 + RPB1 + RPB2 3204aa連結による節足動物の系統樹(ML-GA, α=0.21)。枝上にある数字は信頼度を表している。
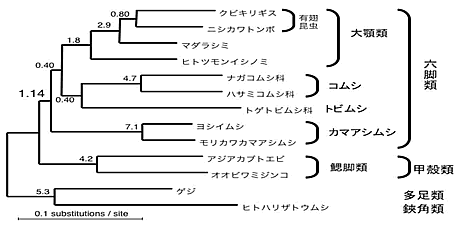 |
図6. DPD1 + RPB1 + RPB2 3412aa 連結 による節足動物の系統樹(ML-GA, α=0.21)。