年度別活動報告
年度別活動報告書:2005年度
3-1.昆虫と植物との共生関係、共進化および種分化に関する研究
蘇 智慧(研究員、代表者)
佐々木剛(奨励研究員)
石渡啓介(大学院生)
神田嗣子(研究補助員)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士或いは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物はそれぞれ陸上で最も多様化した生物群で、その多様化は植物と昆虫とが互いに相互適応的関係を築くことによって促されてきた。したがって、植物と昆虫との相互関係(競争、共生、寄生など)を解明することは、生物の多様性を理解する上で最も重要なカギの一つである。我々は現在植物と昆虫の共生関係の中で最も代表的な系といわれるイチジク属とイチジクコバチの相利共生関係の構築、維持、崩壊のメカニズム、種分化や多様化に与える影響などについてDNA系統解析や野外観察調査などにより研究を行っている。
イチジク属Ficusは、クワ科 (Moraceae) に属し、4亜属 (イチジク亜属Ficus, アコウ亜属Urostigma, Pharmacosycea, Sycomorus)に分けられ、東南アジア、アフリカ、中南米など熱帯を中心に約750種が世界中に分布している。日本はイチジク属の分布域の北限にあたり、南西諸島を中心にアコウ亜属3種、イチジク亜属13種が分布している。そのうち、小笠原諸島に固有種3種と移入種1種がいる。一方、イチジク属植物に共生、或いは寄生している一群のイチジクコバチの仲間は、分類学的に膜翅目のコバチ上科Chalcidoideaに属し、イチジクコバチ科Agaonidaeとされている。イチジクコバチ科はさらに複数の亜科に分けられ、その内の1つが送粉コバチ亜科Agaoninae (fig-pollinating wasps)で、他の亜科はすべて非送粉コバチによって構成されている。
イチジク属植物と送粉コバチとの「1種対1種」の関係は古くから共進化のモデル系として多くの研究がなされてきた。日本産イチジク属とイチジクコバチについても研究が行われている 1)。特に近年アフリカ産を中心に分子系統解析の研究も盛んに行われるようになった。これまでの報告を総合すると、イチジク属植物と送粉コバチの関係は、まだ解明されていない部分もあるが、基本的には両者が共進化してきたと考えられている。しかし、いくつかの問題点も残っていると私たちは考えている。イチジク属の各種がそれぞれ広い分布域を持っており、日本産の大部分の種も東南アジア、オーストラリアまで広く分布している。これほど広い分布域をもつ生物種の系統関係を調べるには、ある程度地域性も考慮しなければならない。特に近縁種間の系統関係を解析する場合、1サンプルだけで種の系統的位置を決めるのはかなり危険であり、異なった結論に導く可能性がある。また、「1種対1種」の関係を検証するためには、分布域を網羅する複数地点からの送粉コバチの材料を分析するのはなおさら重要である。しかし、これまでの報告を見ると、ほとんどの系統解析には1種ごとに1サンプルしか用いていない。もう一つの問題点は系統解析そのものの信頼度である。イチジク属植物の系統解析は核ITS配列を使うのがほとんどである。近縁種間の系統関係はある程度判明しているが、高次分類群の系統関係はほとんど明らかになっていない。結局、形態形質を加えて、分子情報と形態形質を結合した解析を行っている 2)。しかし、全く進化原理の異なる情報を混ぜて正しい系統樹ができるであろうかという疑問も生じる。イチジクコバチの系統解析にも同様な問題がある 3)。
我々は以上の問題点を考慮した上、これまでメキシコ産、日本産と中国海南島産のイチジクとイチジクコバチの分子系統解析を行ってきた。メキシコ産の材料を分析したところ、Urostigma亜属の送粉コバチはおよそ2100万年前に一斉放散分化したことが示唆され、また、同じ種のイチジクから採集した送粉コバチが系統樹上1つの枝にまとまらず、異なる系統に分かれるという興味深い結果が得られた4)。この結果はイチジク属植物と送粉コバチの「1種対1種」関係の崩壊を示唆した。とことが、日本産の材料を調べたところ、その「1種対1種」の関係はほぼ厳密に維持されていることが分かった5), 6)。イチジク属植物と送粉コバチの系統関係について、これまで近縁種間の関係がほぼ解明されたが、上位群間の関係はまだ解明に至っていない。今年度は目的に合わせて異なった分子マーカーを使用したり、新しい分子マーカーを追加したりすることによって、上位分類群間の系統関係の解明に努めた。
結果と考察
1) 日本産イチジク属植物の系統解析
植物は遺伝子の進化速度が遅いため、昆虫に比べると系統解析が難しい。我々は、近縁種間の系統関係と高次分類群の関係を分けて、別々の分子マーカーを使うことにした。前者に対しては核ITS領域を使用し、後者には葉緑体DNAの6つのイントロン部分の塩基配列を用いた。ITSは核ゲノム上に、複数コピーがあるので、クローニングして、1サンプルに対して5-10クローンを取って塩基配列の決定を行った。全DNAは乾燥葉から抽出し、PCR反応と塩基配列の決定を行った。系統解析は、MAFFTでアライメントし、PAUPとMOLPHYでNJとMLの系統樹を作成した。
ITSの配列にはクローン間の違いが多少見られるが、種間関係を越えるほどのものではなく、各種が綺麗にまとまった。但し、小笠原諸島固有種のトキワイヌビワFicusboninshimaeとオオトキイヌビワF. nishimuraeおよび本州と南西諸島に分布するイヌビワF. erectaの3種の間には、有意な違いが見られなかった。葉緑体DNAにおいても3種は全く同じ配列を示した。この結果から、これら3種は非常に近縁であることが示唆された。また、ヒメイタビF. thunbergiiとイタビカズラF. nipponica両種は、葉緑体DNAにおいても、ITS配列においても、同じ配列であり、この2種も極めて近縁であることが判明した。ITSの系統樹では、ガジュマルF. microcarpaとアコウF. superbaはそれぞれ独立した系統を形成したが、他の種類は3つのグループに綺麗にまとまり、グループ内の種間関係も明らかとなった。しかし、グループ間の関係は不明瞭であった(図1)。一方、葉緑体DNAの系統樹はITSの結果を補うように、近縁種間の関係はあまり明瞭ではなかったが、上位群の系統関係は綺麗に判明した。特に雌雄同株のアコウとガジュマルはクラスターを形成することが分かった(図2)。両系統樹の結果を総合して作成した簡略系統図を図3に示した。今回の解析によって、日本産イチジク属の各種の系統関係はほぼ完璧に解明されたと思われる。ちなみに、小笠原諸島のオオヤマイチジクF. iidaianaはフィールドでの採集はできなかったため、解析に加えていないが、その後、乾燥標本を用いて一部の葉緑体DNA配列を解析したところ、小笠原諸島にある他の2種と違いがなかった。つまり、3固有種の違いは塩基配列のレベルでは認められないと結論できる。
2) 日本産イチジク送粉コバチの系統解析
コバチの系統解析は、これまで核28S rRNA遺伝子とミトコンドリアCOI遺伝子を用いて行ってきた。両系統樹において共通して認められた3つのグループは以下の通りである。1)ガジュマルコバチ/アコウコバチ;2)オオバイヌビワコバチ/アカメイヌビワコバチ/ギランイヌビワコバチ;3)ホソバムクイヌビワコバチ/ムクイヌビワコバチ/ハマイヌビワコバチ。これら3グループ内の関係も一致していた。しかし、グループ間の系統関係は、どちらにおいても高い支持率が得られず、解明には至らなかった。これら上位分類群間の関係を明らかするために、今回はND1,ND4,ND5,Cytbなどのミトコンドリア遺伝子の追加解析を行ったが、種によってまだ一部の配列しか得られていないため、最終的な系統解析はまだできない。途中の解析の結果、アコウコバチとガジュマルコバチが近縁関係を示し、他の送粉コバチと最初に分岐することが示唆された。宿主のイチジク属植物の系統関係と比較してみたところ、解明された部分は基本的に一致していると判明した(図4)。解明に至っていない部分の中で、例えばイヌビワとオオイタビは近縁関係を示しているが、それらの送粉コバチは明確に組んでいない。しかし、両種は他のコバチとの関係もはっきりしていないので、共通祖先由来ではないとまでは言えない。最終的な結論は送粉コバチの上位関係の完全解明を待たなければならないが、これまでの結果を総合すると、日本産イチジク属植物とそれらの送粉コバチは基本的には同調的に種分化(cospeciation)してきたと考えてよいだろう。
3) 「1種対1種」関係の厳密性と例外発見
イチジク属植物と送粉コバチの「1種対1種」関係について、これまでの解析結果から、相当厳密に維持していることがはっきりした。同種のイチジク属植物から採集された送粉コバチは、系統樹上乱れることなく、同じ配列を示すか、僅かな違いがあるにしても綺麗にまとまっていた。しかし、興味深い例外が見つかっている。ギランイヌビワの花嚢から採集した送粉コバチはアカメイヌビワコバチと全く同じ配列を示し、またアカメイヌビワとイヌビワから採集した送粉コバチが、逆にギランイヌビワコバチと同配列だったのである。これは、アカメイヌビワコバチがギランイヌビワの花嚢に進入でき、逆にギランイヌビワコバチがアカメイヌビワとイヌビワの花嚢に進入できることを意味する。言い換えれば、送粉コバチはある環境下(例えば、本来の宿主種がまわりにいない)において、別種の花嚢にも入ることが可能である柔軟性を持っていることが示唆された。但し、これらのコバチはすべて親コバチであるため、産卵できたかどうか、子どもコバチができたかどうかは確認されていない。
4) 小笠原諸島の固有種と移入種の起源
前にも述べたように、小笠原諸島には固有種3種(トキワイヌビワ、オオトキワイヌビワとオオヤマイチジク)と移入種1種(ガジュマル)が分布している。オオヤマイチジクを除いた他の3種の材料は採集することができた。解析の結果によると、トキワイヌビワとオオトキワイヌビワは全く区別がつかないほど近縁なものであり、それらの送粉コバチも同様差異はなかった。また、これら2種は、植物も送粉コバチも、それぞれ本州と南西諸島に広く分布しているイヌビワおよびその送粉コバチと近縁であることが分かった。オオヤマイチジクについては、植物乾燥標本を用いた一部の解析結果から、トキワイヌビワ、オオトキワイヌビワとイヌビワに極めて近縁であることが示唆された。これはYokoyama 1) の結果とも一致する。以上のことから、小笠原諸島の固有種3種は、南西諸島(或いは本州)から進入したイヌビワが島内で種分化したものであると考えられる。但し、トキワイヌビワとオオトキワイヌビワは本当に2種にすべきか、再検討する必要がある。
移入種のガジュマルは明治初期に防風林として移入・栽培されたものである。長い間、結実していなかったが、近年実が付くようになったと言われている。換言すれば、ガジュマルは近年になってようやく受粉できるようになったのである。その送粉コバチの正体は興味深い。
トキワイヌビワなど3固有種の送粉コバチがガジュマルにも送粉するようになったのか(宿主の転換)、或いは本来のガジュマルコバチが近年移入されたのかを調べたところ、小笠原のガジュマルコバチは南西諸島のものと全く同じであることが分かった。つまり、もし起きていれば面白いと思った宿主転換ではなく、人為的にガジュマルコバチが近年になって移入されたのだろう。この事実からも、イチジクとイチジクコバチの「1種対1種」関係は非常に堅いものと思われる。
おわりに
イチジク属とイチジクコバチの関係は寄生コバチを含めた三者の関係が絡み合った複雑な系である。イチジク属と送粉コバチの共生関係の進化を探る上ではこれら三者の生活史がどのように関連しているかを知る必要があるが、まだほとんど分かっていないのが現状である。今後は、送粉コバチの上位分類群の系統関係解明、非送粉コバチの詳しい系統関係、中国産イチジク属植物とイチジクコバチの系統関係および日本産のものとの繋がりなどを中心に解析を行い、イチジクコバチによる宿主転換の可能性や転換機構および種分化に与える影響などに関する理解を深めたい。
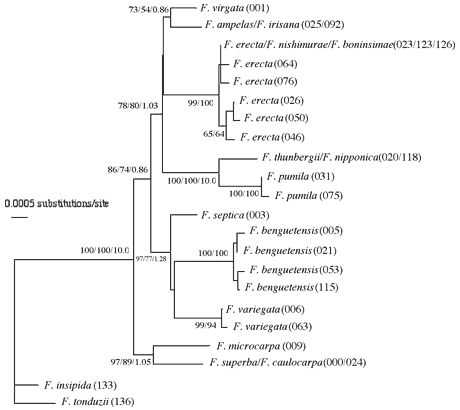 |
図1. 葉緑体DNAによる日本産イチジク属植物の系統樹。枝の分岐点にある数字はBootstrap確率など系統樹の信頼度を示している。左からNJ (PAUP), ML(PAUP)およびML(MOLPHY)。
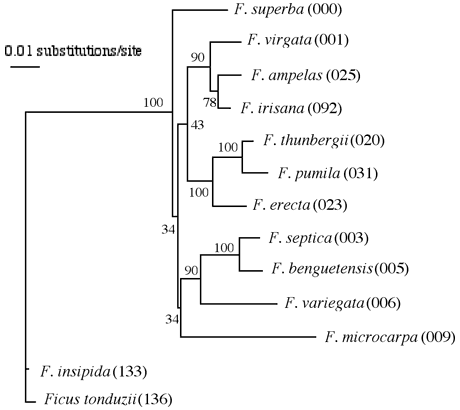 |
図2. ITS配列によるイチジク属植物の系統樹。PAUPで作成したML系統樹である。枝上の数字は信頼度を表すBootstrap確率である。
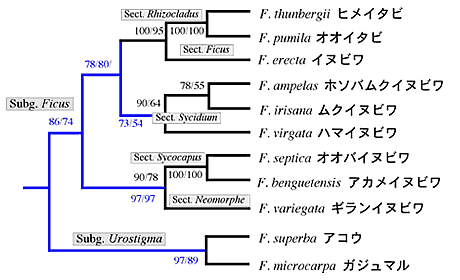 |
図3. 葉緑体DNAと核ITS領域によって解明された日本産イチジク属植物の系統関係。上位関係は葉緑体の系統樹(青枝)を用い、近縁関係はITSの系統樹(黒枝)を用いた。Bootstrap確率は左からNJおよびMLのものである。分類学的な亜属とセクションは系統図に示した。
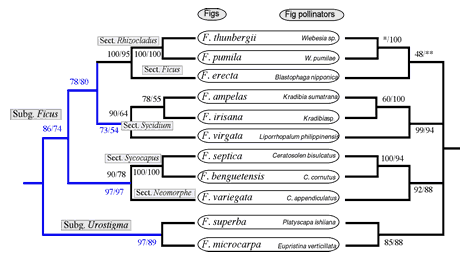 |
図4. 日本産イチジク属植物の系統図(左)とそれらの送粉コバチの系統図(右)の対応関係。枝の上の数字は系統樹の信頼度を表すBootstrap値である。植物の系統関係は、ほぼ完璧に解明されたが、送粉コバチの上位分類群の系統関係はまだ解明に至っていない。