年度別活動報告
年度別活動報告書:2013年度
分子系統から生物進化を探る 3-1.昆虫と植物との共生・共進化を介した種分化機構の解明
蘇 智慧(主任研究員) 和智仲是(奨励研究員)
佐々木綾子(研究補助員) 宮澤秀幸(大阪大学大学院生)
はじめに
様々な生物種から構成される地球生態系のなかでの生物同士あるいは生物と環境との相互作用は、生物の多様性を生み出す大きな原動力と考えられる。昆虫と被子植物はそれぞれ陸上で最も多様化した生物群で、その多様化は互いに相互適応的関係を築くことによって促されてきたように見える。その最たる例が「1種対1種」の密接な送粉共生関係を築くイチジク属植物 (クワ科) と送粉コバチ(イチジクコバチ科、以下イチジクコバチ)であり、双方が750種以上の種数を誇っている。我々は現在イチジク属植物とイチジクコバチとの共生関係の構築過程と維持機構およびその共生系における種分化機構などについて研究を行っている。
イチジク属植物は、東南アジア・アフリカ・中南米など熱帯を中心に分布している。日本はイチジク属植物の分布域の北限にあたり、南西諸島を中心に16種が生息している。イチジク属植物は花嚢とよばれる袋状の閉じた花序をつけ、その内側に多数の花を咲かせる。送粉者であるイチジクコバチは、この花嚢に入り花粉を媒介するだけでなく同時に花に産卵する。ふ化した幼虫は虫こぶ(昆虫が改変した植物の組織)の中で成長する。やがて次世代のイチジクコバチが花嚢内で羽化・交配し、雌成虫が花粉を持って他の花嚢へと移動することで次の受粉が成立する。このようにイチジク属植物とイチジクコバチの二者は、繁殖において互いに強く依存し合った関係と言える。
イチジク属植物とイチジクコバチとの共生関係は、「1種対1種」という種特異性が極めて高いものと言われている。この「1種対1種」関係を維持しながら種分化が起きるとしたら、同調した種分化や系統分化が起こることが予想される。これまで分子系統学的解析を用いてこの仮説を検証する研究が行われてきた。その結果、イチジク属植物の節(植物では属・亜属・節・種と分類する)レベルの系統関係とイチジクコバチの属レベルの系統関係がおおまかに一致し、仮説が支持されている。しかし種間・種内レベルでの系統関係の矛盾のほかに、イチジクコバチの隠蔽種や1種のイチジク属植物に複数種のイチジクコバチ(あるいはその逆)が共生するなど、不明瞭な点が残っている1-4)。我々のこれまでの研究によって、日本産のイチジク属植物とイチジクコバチでは「1種対1種」関係の厳密性が見られ、同調的系統分化が示唆された5)。一方でメキシコ産の イチジク属植物とイチジクコバチでは、近縁種間で「1種対1種」関係の乱れが見られた2)。また、小笠原諸島産のイチジク属植物の進化・種分化の過程においては異種間交雑が起きていたことが示唆され、雑種形成がイチジク属植物の種分化をもたらす要因の1つであると考えられた6)。また、イチジク属植物が特定のイチジクコバチを自らの花へ呼び寄せるための手段として、花の匂い(花から放出される分子量300以下の揮発性に富んだ化学物質の集まり)が注目されてきた。多くの送粉者は匂いを嗅覚によって捉え花の探索に利用している。イチジク属植物でも、花嚢の外見は緑色で非常に目立ちにくいため視覚情報よりも嗅覚情報としての花の匂いが有効であると考えられる。
このような背景のもと、本年度は主にイヌビワとその近縁種およびそれぞれのイチジクコバチの種間・種内の関係に注目して、集団遺伝学的解析、花の匂いを認識する遺伝子の探索を行った。
結果と考察
1)イヌビワとその近縁種3種を利用するイチジクコバチの集団遺伝学的解析
イヌビワ(Ficus erecta)は東南アジアから東アジアにかけて広く分布しており、日本・韓国・台湾・中国から知られている。さらに台湾にはイヌビワに近縁な3種F. formosana, F. tannoensisとF. vaccinioides が分布している。これら4種の近縁な植物種に対して、それぞれ種特異的に共生関係にあるイチジクコバチが知られている(表1)。植物種は形態的・遺伝的に明確に区別ができる。その一方で、F. vaccinioidesを利用するBlastophaga yeniを除くイチジクコバチ3種は、形態的によく似ており、記載時に用いられた種の識別形質も連続的で曖昧である。
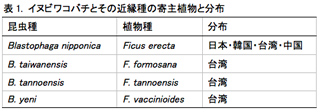
これらの4つの絶対共生系の構築過程・維持機構を理解するために、これまで植物の系統解析・花の匂い物質の解析が行われてきた(2011年度・2012年度活動報告)。その結果、植物種は、遺伝的にも花の匂い物質の組成も分化していることが明らかになった。もし、この4つの絶対共生系でも、他の多くのイチジクコバチで示唆されているように同調的な種分化が起こっているのであれば、コバチにも寄主植物に対応した遺伝的分化が見られるはずである。本年度はそのことを検証するために、コバチを対象に集団遺伝学的解析を行った。
各地から採集した試料の雌個体からDNAを抽出し、ミトコンドリアDNAの部分配列(COX1-COX2, CYTB)、核DNAの13遺伝子座(マイクロサテライトマーカー)を用いて比較解析を行った。解析にはMEGA, GenAlExを用いた。その結果、下記の2点が明らかになった。
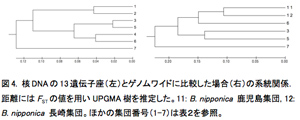
1.1. Blastophaga nipponica, B. taiwanensis, B. tannoensis は寄主植物に対応した遺伝的分化が見られない.ミトコンドリアDNAから推定された系統関係(図1)からも、核DNAの13遺伝子座から推定された遺伝的分化(表2, 図2)からも、台湾に分布する3種(B. nipponica, B. taiwanensis, B. tannoensis)を明確に区別することはできなかった。このことは3種が実は形態的にも遺伝的にも分化していない同一種である可能性を示唆している。このような、1種(もしくは極めて近縁種)のコバチが複数種の植物を利用している例は世界的にもまれである。今後、形態的・遺伝的分化についてさらに精査を行っていく予定である。
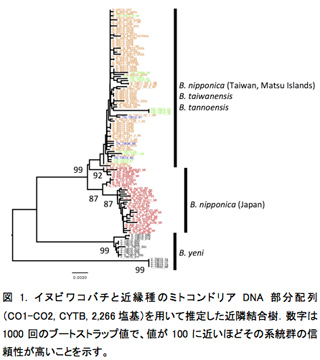
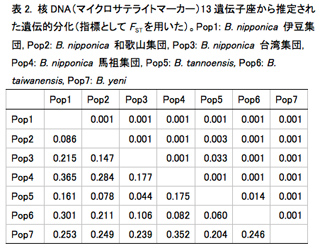
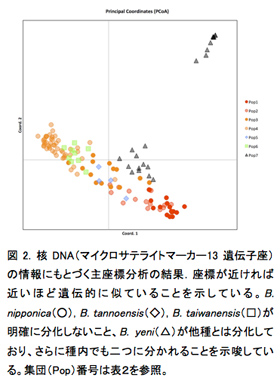
1.2. Blastophaga yeni は他の近縁種と遺伝的にも区別できる一方、種内にも明確な分集団が見られる.ミトコンドリアDNAでも核DNAでも、他3種とは異なり、B. yeni には寄主植物に対応した遺伝的分化が見られた(図1、表2)。意外なことに、種内にも分化が見られ、2つに分化することが明らかになった(図1, 2)。台湾に広く分布するイチジクコバチCeratosolen bisulcatus(2012年度活動報告)やWiebesia pumilaeは明確な分集団化は見られない一方で、台湾南東部の狭い範囲にしか分布していないB. yeni は分集団化が起こっていることが興味深い。
2)イヌビワコバチのゲノム解読の試み
イヌビワコバチの分子系統的・集団遺伝学的研究を効率的に進める上で、ゲノムに関する情報の取得と解析は欠かせない。イヌビワコバチに比較的近縁なキョウソヤドリコバチNasonia vitripennisのゲノム情報は公開されているが、より流用性が高いと考えられるイチジクコバチ科昆虫のゲノム解読は未だ行われていない。そこで、より効率的に研究を展開するために、次世代シーケンサ(Ilumina, Miseq)を用い、イヌビワコバチのゲノム解読を試みた。無作為に断片化したゲノムDNAから構築したライブラリーを用い、2ラン分の情報を取得した。その結果、およそ3,500万の塩基配列が得られた。CLC Genomic Workbenchを用いて、これらの配列のアセンブル(相同な配列をまとめること)を行った結果、最大で84,000塩基のコンティグ配列(ゲノム上のひとつながりの塩基配列)が78,000個得られた(表3)。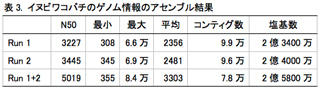
さらに相同性検索などを行えば、分子系統的・集団遺伝学的研究に用いることのできる新規の中立マーカーの開発等に、今回得られた情報が有効に活用できることが期待される。全ゲノム情報を網羅するには十分とは言えないため、今後さらに解析を行い、より網羅性の高い充実したゲノム情報にできればと考えている。
3)イヌビワコバチとその近縁種を用いた寄主植物適応遺伝子の探索
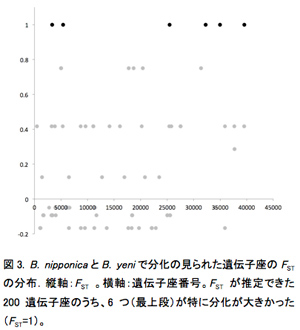
イチジク属植物には基本的に種特異的共生関係にあるイチジクコバチが知られている。それぞれのコバチの寄主植物に対する特異性は具体的にコバチのゲノムのどのような違いに起因しているのだろうか。この疑問に答えるために、イヌビワコバチとその近縁種(1参照)を用いて、寄主植物適応遺伝子の探索を試みた。ゲノムの相同な部分を網羅的に調べるために、制限酵素でゲノムを選択的に切断し、ライブラリーを作成する方法(ddRAD-seq)を用いた。8〜10個体の雌個体から得られたゲノムDNAをプールしたライブラリーを作成し、次世代シーケンサにより配列情報を取得した。得られた情報を解析し、種特異的・寄主植物特異的な点突然変異を探索した。解析には Stacksを用いた。B. nipponica(日本集団・台湾集団), B. tannoensis, B. taiwaensis, B. yeni を解析した結果、19,259遺伝子座(ゲノム上の相同な位置にある断片)の情報が得られた。もし、寄主植物に適応したゲノム領域があるとすれば、寄主植物の異なるB. nipponicaとB. yeniを比較したときにその領域の分化が大きいことが予想される。予備的な解析の結果では、そのような遺伝子座は6つ検出された(図3)。これらの遺伝子座の近くに寄主植物に適応したゲノム領域がある可能性がある。今後、同様の解析を他の種・集団について種間で分化の大きな遺伝子座の検出を試みる予定である。しかし複数個体のゲノムDNAをプールする方法は統計的な検出力が落ちたり、サンプリングの偏りによる誤った結果を導いたりする可能性が指摘されている。実際、上記のデータにおいて、遺伝子座として検出する閾値を上げると、遺伝子座の数は8,795と減少した。さらにB. nipponicaとB. yeniを比較したときに分化が大きい遺伝子座は見られなくなった。今回の結果も人為的産物である可能性があるため、今後さらに個体ごとに識別して解析する実験系を確立し、広く分布域をカバーした試料を用いた再解析を行う予定である。
また本来の目的ではないが、これらの遺伝子座の配列情報をもとに、MEGAを用いて4種の系統関係を推定したところ、これまでミトコンドリアDNAや核DNAで推定されている系統関係(1参照)とほぼ矛盾しない結果になった(図1, 4)。ゲノムワイドに比較した結果でも、台湾のB. nipponica, B. tannoensis, B. taiwanensis が明確に分化しないことが示唆された(1参照)。
4)イヌビワコバチとその近縁種の寄主植物認識遺伝子の探索
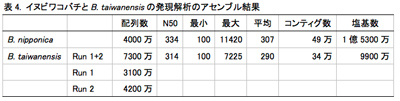
イチジクコバチはそれぞれ特異的なイチジク属植物を利用するために形態的にも生態的にも適応しているように見える。そして、寄主植物種を認識するのには嗅覚情報を利用しているのではないかと考えられている。しかし、イチジクコバチにおいて、寄主植物認識にどのような遺伝子が重要な役割を担っているかという具体的な機構は十分には明らかになっていない。この寄主植物認識遺伝子を明らかにすることができれば、イチジクコバチがどのようにして寄主植物を認識し、どうやって適応してきたかという共生の進化についての重要な知見が得られることが期待される。また、イチジクコバチの種分化機構の解明にも繋がる。そこで、イヌビワコバチとその近縁種(1参照)の頭部で発現している遺伝子群を網羅的に解析することにより、主に嗅覚情報の認識に関与している可能性のある遺伝子の探索・同定を試みた。イヌビワコバチの雌の生体頭部からRNAを抽出し、cDNAを合成した。またB. taiwanensis については野外で採集した個体をRNA保存液(RNAlater)に浸したものを用いた。RNAから合成したcDNAをライブラリー化し、次世代シーケンサで解析を行った。得られた配列データの解析には CLC Genomic Workbench、blast2go、自作のpythonスクリプトを用いた。その結果、イヌビワコバチでは4000万、B. taiwanensisでは7400万の配列情報が得られ、それぞれ50万、34万のコンティグ配列が得られた(表4)。
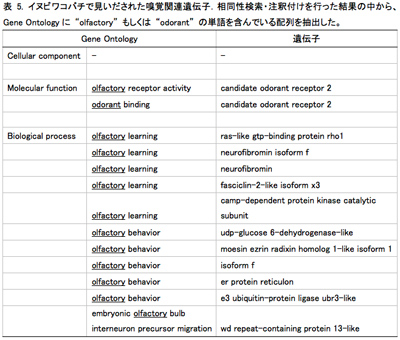
得られた配列情報の一部をもとに、予備的解析として相同性検索・注釈付けを行った(図5)ところ、イヌビワコバチでは、10種類の嗅覚に関係すると思われる遺伝子配列が見いだされた(表5)。このうちOlfactory receptor 2(Or2)はイチジクコバチの一種 Ceratosoren solmsi で同定された寄主植物認識の関与が示唆されている遺伝子で、多くの昆虫種で保存されているOrco の相同遺伝子である。今後、さらに解析を進め、嗅覚関連遺伝子の探索を行うとともに、見いだされた遺伝子のDNA配列の種間・種内の遺伝的変異を調べる予定である。
一方で B. taiwanensis では嗅覚関連遺伝子の配列は見つからなかった。またコンティグ配列の長さや相同性検索・注釈付けの結果から、今回用いたB. taiwanensis 試料はRNAが劣化している可能性が高いことが示唆された。今後、イヌビワコバチを含め近縁種のRNA発現解析に耐え得る試料の保存方法の検討を行っていく予定である。

おわりに
イチジク属とイチジクコバチとの共生・共種分化機構を解明するためには、これまで主として分子系統解析と集団遺伝学的解析の手法を用いて行ってきた。これらの解析に加え、3年前から花の匂いの比較解析も行ってきた。本年度は主にイヌビワとその近縁種およびそれぞれのイチジクコバチの種間・種内の関係に注目して、集団遺伝学的解析・花の匂いを認識する遺伝子の探索を行った。その結果、イヌビワと近縁種2種にそれぞれ送粉している「3種」のイチジクコバチは遺伝的分化が全く見られないことが判明した。このことは3種が実は形態的にも遺伝的にも分化していない同一種である可能性を示唆している。このような、1種のコバチが複数種の植物を利用している「多種対1種」の例は世界的にもまれであり、今後、形態的・遺伝的分化についてさらに精査を行っていく予定である。また、今年度は寄主植物適応遺伝子の探索と花の匂いを認識する遺伝子の探索を試みた。まだ確実な成果をあげていないが、今後の研究を進めていくうえで有益な多くの知見が得られた。これらの新たな取り組みをイチジクとイチジクコバチの種分化機構の解明に繋げて行きたい。