年度別活動報告
年度別活動報告書:2006年度
3.分子系統から生物進化を探る 3-2.昆虫類(六脚類)の起源、系統と進化に関する研究
蘇 智慧(研究員、代表者)
佐々木剛(奨励研究員)
神田嗣子(研究補助員)
石渡啓介(大阪大学大学院生)
はじめに
陸上昆虫類(六脚類 Hexapoda)は、無翅昆虫類と有翅昆虫類とに大きく分けられる。無翅昆虫類は5目(カマアシムシ目、トビムシ目、コムシ目、イシノミ目、シミ目)、有翅昆虫類は27目を含んでいる。形態学による分類では、カマアシムシ目、トビムシ目とコムシ目は内顎類、イシノミ目/シミ目とすべての有翅昆虫は外顎類とされている。従来、六脚類は他の節足動物のうち多足類 (Myriapoda)と近縁であり、六脚類内部では無翅昆虫類から有翅昆虫類のグループが分岐し翅を獲得したと考えられてきたが、最近の分子系統解析により、六脚類は多足類ではなく甲殻類 (Crustaceae) と近縁であることが明らかになった。さらに六脚類の単系統性にも疑問がもたれ、Nardi ら 6) によるミトコンドリア遺伝子を用いた研究では、無翅昆虫類のトビムシ目は甲殻類よりも以前に分岐した可能性が示唆された。しかし、この結果は解析方法によって変わるなど有意なものではなく、また、トビムシ目以外の無翅昆虫類が含まれていないため、六脚類の起源に関する結論には至っていない。Regier ら7)は、核にコードされたタンパク遺伝子RNA polymerase II largest subunit (RPB1), elongation factor 1α (EF-1α), elongation factor 2 (EF-2)を用いた解析により六脚類の単系統性を示したが、EF-1α 遺伝子を用いるのには問題がある(後述)。また、彼らのデータにも無翅昆虫類のカマアシムシ目が含まれていないため、六脚類の単系統性には疑問が残っている。
そこで私たちは多数の核ゲノム上のタンパクをコードしている遺伝子を比較して無翅昆虫類の正しい系統的位置を解明し、昆虫類の起源、系統と進化を明らかにし、六脚類の共通祖先が一度だけ陸上へ進出したのか、それとも複数の系統の祖先が独立に複数回上陸したのかという問いに答えることを目的として、本研究を行っている。
結果と考察
1) 系統解析に適した遺伝子
これまでに、以下3つの条件に基づいて節足動物の系統解析に適した遺伝子の探索を行ってきた。1) 節足動物の系統内部で遺伝子重複がない;2) 適度な保存性を有しており、縮退プライマーの設計が可能である;3) PCRの結果が良好である。18S ribosomal RNA (18S rRNA), Elongation Factor 1 alpha (EF-1α) および 2 (EF-2), RNA polymerase II largest subunit (RPB1), RNA polymerase II second largest subunit (RPB2), DNA polymerase delta catalytic subunit (DPD1), ミトコンドリアゲノム遺伝子などについて検討を行った。その結果、18S rRNA 遺伝子は、データは豊富にあるが、系統間の進化速度のばらつきが大きく、系統解析には向かない。ミトコンドリア遺伝子は塩基置換が飽和している可能性があり、全文にすでに述べたように信頼出来るような系統樹を構築することはできなかった。EF-1α 遺伝子は、六脚類において遺伝子重複が観察され、パラロガスな比較をしてしまう危険性があるため六脚類の系統関係を推定するのに用いることはできない。これらの遺伝子に対して、RPB1, RPB2とDPD1はおおむね上述3つの条件を満たしている。そこで、我々はこれまでに主としてこれら3つの核内遺伝子を用いて系統解析を進めてきた。さらに、この3つの遺伝子に加えて、DNA polymerase zeta catalytic subunit (DPLZ) 遺伝子も検討し、系統解析に適していることがわかり、解析を開始した。
2) 六脚類の系統関係
本研究の遂行に当たって、材料の収集はもっとも重要な作業である。これまで多くの専門家やアマチュアの方々の多大な協力を得て無翅昆虫類、有翅昆虫類、そして甲殻類や他の節足動物の材料を入手した。解析方法は下記の通りである。RNAを抽出して逆転写を行い、cDNA を鋳型にして縮退プライマーを用いたPCR反応を行い、塩基配列決定を行っている。また、RACE法により可能な限り各遺伝子のアライメント可能な領域全長配列を決定している。得られた配列データは複数台の並列コンピューターを用いて、最尤法による分子系統解析を行った。
最尤法に基づく分子系統解析によって昨年度までに、無翅昆虫類のなかでもイシノミ目とシミ目は有翅昆虫類と近縁となり、外顎類(有翅昆虫類+イシノミ目/シミ目)をひとつの分類群とする考え方が強く支持された。しかし、イシノミ目とシミ目は単系統とはならず、有翅昆虫類とシミが近縁でありイシノミはより古くに分岐したことが支持された。内顎類の各グループ(カマアシムシ目、トビムシ目、コムシ目)の分岐順序ははっきりしなかった。有翅昆虫類内部では、トンボとカゲロウの旧翅類が他の新翅類より古くに分岐したことが支持された。
本年度は、内顎類のうち進化速度が大きく、LBA (long branch attraction artifact) の原因になる可能性の大きいオオフォルソムトビムシに代えて、キボシマルトビムシを用いた。さらに、甲殻類十脚目のアメリカザリガニを加え、解析を進めた。最尤法による系統解析の結果、統計的に有意ではないが六脚類の単系統性が示唆された。また、内顎類のカマアシムシ目が、六脚類の中でもっとも古く分岐したことも示唆された(図7)。
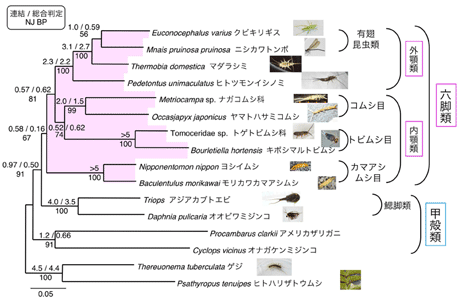 |
図7. DPD1+RPB1+RPB2 3204aaによる節足動物の系統樹(ML-GA WAG-Γ, α=0.21)。枝上にある数字は信頼度を表している。
有翅昆虫内部の系統関係についても解析を進めた。有翅昆虫のなかでトンボ目とカゲロウ目(旧翅類)が最初に分岐し、新翅類が単系統となることが強く支持された。また、従来新翅類の中では、完全変態類と準新翅類が近縁であると考えられてきたが、最尤法による解析の結果、完全変態類は多新翅類と近縁であることが支持された。準新翅類の単系統性は支持されなかった。また、統計的に有意ではないが、完全変態類、多新翅類、そして旧翅類はそれぞれ単系統であることが支持された。(図8)
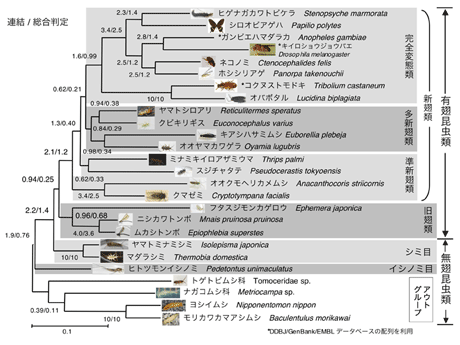 |
図8. DPD1+RPB1+RPB2 3368aa による有翅昆虫類の系統樹 (ML-GA JTT-Γ, α=0.17)。
おわりに
六脚類は単系統(単一起源)なのか?彼らの上陸は一回のみか、それとも二回以上なのか?これは本研究において明らかにさせたい問題である。しかし、これまでに我々は複数の核内遺伝子(約4000アミノ酸残基)を用いて系統解析を行ったところ、陸上昆虫は単系統である可能性が高いことが示唆されたものの、統計的に有意な支持がまだ得られていない。この結果から、甲殻類と昆虫類の分岐および昆虫類の初期種分化は短期間のうちに起きたのではないかとの可能性がますます強まってきたと思われる。もしそうであれば、この問題の解決は極めて困難になる。但し、理論的には分子情報をもっと増やせば有意な結果が得られる可能性も残っている。従って、今後はDPLZ 遺伝子を含め、更なる遺伝子情報を増やし、信頼性の高い系統樹の構築に努めたい。