年度別活動報告
年度別活動報告書:2004年度
アゲハチョウの食草選択と進化
吉川 寛(顧問)
尾崎克久(研究員)
中山忠宣(奨励研究員)
広崎由利恵(研究補助員)
はじめに
昆虫は生活史の中で、環境の情報を知る手段として化合物を巧みに利用している。化学受容は一般に味覚・嗅覚として知られており、寄主選択、配偶行動、集団生活の維持など様々な場面で重要な役割を持っている。寄主選択において化学受容の仕組みに変化が生じた場合、それまでとは異なる空間を生息の場として利用する集団が現れ、住み分けが何世代にもわたって繰り返されることによって種分化へとつながる。昆虫と環境との関わりにおいて中心的な機能である味覚や嗅覚といった化学受容に関わる分子の解明は、昆虫の多様化、種分化、進化の仕組みを解明するために、最も有力な手がかりになると考えられる。
アゲハチョウの仲間は単食性に近い寄主選択をしており、寄主選択とアゲハチョウ科の進化には相関関係が認められる。このことに注目して、雌成虫が前脚ふ節に存在する感覚毛で食草に含まれる化合物を認識し、産卵行動が刺激されることが明らかにされた。最も良く研究されているナミアゲハ(Papilio xuthus)では、ウンシュウミカンの葉から産卵刺激物質として10種類の化合物が単離されている1)。それらは、主としてフラボノイド、アミン類、糖の誘導体に分類される。他にも数種のアゲハ類で産卵刺激物質が明らかにされており、これらの構造についてアゲハ種間で比較すると類似性が認められる。つまり、植物の系統的近縁性とは無関係に、植物に含まれている化合物の類似性が寄主転換の可能性を支え、食性の進化を可能にしたのではないかと考えられている2)。産卵刺激物質を受容する感覚毛は、ショウジョウバエの典型的な味覚組織に構造が類似し、味覚細胞が存在することから、産卵刺激物質特異的な味覚レセプターを発現していることが予測される。アゲハチョウの仲間が産卵刺激物質として認識する化合物の組み合わせは種ごとに異なるため、味覚レセプターは種ごとに特徴的な多様なレパートリーを持っていることが予想される。それらを網羅的に解析し、特徴を比較することで、レセプターの進化と食性の変化の関係を解き、種分化の謎に迫れると考えた。
昆虫の味覚レセプターの研究は2000年になって初めてショウジョウバエから候補遺伝子として新規の7回膜貫通Gタンパク質共役型レセプター(以下GPCRと略)ファミリーが同定された3)。以降、ハマダラカとオオタバコガについて化学感覚器官に特異的に発現するGPCRが発見されている4) 5)。これらのレセプターは構造が多様であるため、類似性による探索は困難で、いずれも全ゲノム配列情報の情報科学的解析から明らかにされている。また、味覚、嗅覚などの化合物がリンパ液中の細胞表面に存在するレセプターに到達するために、これら化合物を結合し運搬するタンパク(Chemosensory protein=CSP、Odorant Binding Protein=OBP)が知られている。これについても、ショウジョウバエ全ゲノムの解析から昆虫には多種類のCSP・OBP遺伝子が存在し、味覚器官に特異的に発現するものも見つかっている6)。このように昆虫の味覚レセプターとCSP・OBPに関する配列情報が蓄積されつつあることにより、情報科学的手法を用いてアゲハチョウの産卵刺激物質認識に関わる遺伝子群をクローニングする道が開けた。
そこで、主たる食草の産卵刺激物質が明らかにされているナミアゲハを材料として用い、雌成虫前脚ふ節に発現する味覚レセプター遺伝子及びCSP・OBP遺伝子をクローニングし、その機能と特徴の解明を目的とした。初年度(2001年)はアゲハ雌成虫前脚ふ節からcDNAライブラリーを調製し、大量塩基配列決定方法とRT-PCRによる発現部位決定法を確立した。2002年度はcDNAクローンの網羅的な塩基配列決定及び、得られた配列の情報解析、発現解析を行った。その結果、雌成虫ふ節に特異的に発現するGPCRであるPXFT-01989、雌雄の化学感覚器官に優先的に発現する3種類のCSP、雌成虫ふ節で発現する3種のアミンレセプター及び1種類のチトクロームp450の遺伝子をクローニングした7)。2003年度はFosmidベクターを用いてナミアゲハゲノムライブラリーを作製し、既に得られた上記候補遺伝子をプローブとして、ゲノム上の遺伝子の全構造と関連遺伝子ファミリーのクローニングを行った。
今年度は、ナミアゲハ雌成虫ふ節に特異的に発現するGPCR遺伝子であるPXFT-01989から、選択的スプライシングによって少なくとも4種類のメッセンジャーRNAが作られていることを発見した。これら転写産物を培養昆虫細胞中で発現させ、カルシウムイメージング法でリガンドの特定を試みた。また、PXFT-01989遺伝子を手がかりとして、他のアゲハチョウからGPCR遺伝子を探索した結果、シロオビアゲハから類似性の高い部分配列を発見した。複数のGPCR遺伝子の発見を目的として、シロオビアゲハ雌成虫ふ節の完全長cDNAライブラリーと、Fosmidベクターを用いたゲノムライブラリーを作成した。
結果と考察
1. GPCR遺伝子PXFT-01989の選択的スプライシング
ゲノム配列にcDNA配列をアライメントした結果、ナミアゲハ雌成虫ふ節に特異的に発現するGPCR遺伝子は、エキソンが8つに分断されていることが明らかになった(図1)。
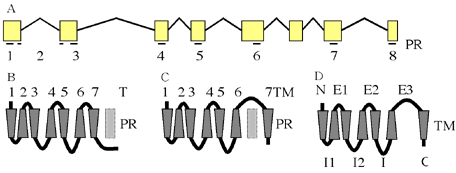 |
図1:PXFT-01989ノーマルタイプの遺伝子構造と推定タンパク構造
A: 遺伝子構造。エクソン部分を矩型、イントロン部分を山形の線で表す。エクソン部分の下にTM-PR を線で示す。B: SOSUIによるTMとループのトボロジーの推定。TM-PRのうち、TMと予測されている場所を台形、TMと予測されなかった場所を薄い四角で示す。C: TMHMMによるTMとループのトボロジーの推定。TM-PRのうち、TMと予測されている場所を台形、TMと予測されなかった場所を薄い四角で示す。AとBでTM7の場所が異なる。D: GPCRにおける各部の名称。N=Nテイル、C=Cテイル、E1=細胞外ループ1、E2=細胞外ループ2、E3=細胞外ループ3、I1=細胞内ループ1、I2=細胞内ループ2、I3=細胞内ループ3。
8個のエキソン全てがアミノ酸コード領域として使われているメッセンジャーRNAが最も発現量が多かったので、これをノーマルタイプと呼ぶ。ノーマルタイプのコード領域は1224塩基(407アミノ酸)である。幕貫通領域推定アルゴリズムを用いてタンパクの構造を予測した結果、基本的に膜貫通領域(TM)はエキソンの中に収まり、例外的に、TM2だけがイントロンによって分断されていた。4種類の膜貫通領域推定アルゴリズム(SOSUI、TMHMM、TMAP、HMMTOP)の解析結果で、TM1からTM6は数残基程度のずれはあるもののほぼ共通であった。TM7の位置は2種類に分かれ、TM6の近くに推定されてE3ループが短くCテイルが長い場合と、C末端の近くに推定されてE3ループが長くCテイルが短い場合があった。代表的なアルゴリズムの名前をとって、前者をSOSUI型、後者をTMHMM型と呼ぶことにする。TM7と推定される場所が二カ所あることから、このタンパクには膜貫通領域になり得る場所(Trans Membrane Potential Region: TM-PR)が8カ所あることが解る。TM-PRをN末端側からC末端側の順番で、PR1からPR8と呼ぶことにする。RT-PCRによる増幅産物を詳細に解析した結果、選択的スプライシングによってノーマルタイプより短縮したメッセンジャーRNAが3種類見つかった(図2)。それぞれをクローニングし、塩基配列を決定したところ、いずれも構造から予測できるスプライトサイトで生じているスプライシング多型であることが判明した(図3)。ショートタイプAは、第1エキソンの後半部分が隣接するイントロンと融合して切り取られるという変則的なスプライシングにより、138塩基(46アミノ酸)短縮していた。切り取られる部分に、PR1の後半部分が含まれている。PR2がイントロンによって分断され、後半部分が第2エキソンに乗っているので、スプライシング後はPR1の前半部分とPR2の後半部分が結合する。ショートタイプBは、第4エキソンが両隣のイントロンとともに切り取られ、153塩基(51アミノ酸)短縮していた。第4エキソンにはPR5が含まれる。ショートタイプCは、第5エキソンが両隣のイントロンとともに切り取られ、213塩基(71アミノ酸)短縮していた。第5エキソンにはPR6が含まれる。各ショートタイプはTM-PRを一つずつ失っているが、TM-PRが7カ所残るので全てが7回膜貫通型になることは可能である。しかし、残っているTM-PRを利用して7回膜貫通型になった場合、部分的に膜貫通領域の向きやループ部分の細胞質側・細胞外側といったトポロジーが逆転することになる。発現量は、ショートタイプ全てを合計したものとノーマルタイプがほぼ同量だった(図2)。この結果から、産卵刺激物質受容体としては、4種類の転写産物のうちノーマルタイプが中心であると考えられる。
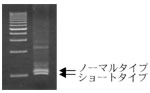 |
図2:-PCR:ーハス?による発現の確認
ノーマルタイプの発現量と、ショートタイプ全種の合計発現量がほぼ同じである。
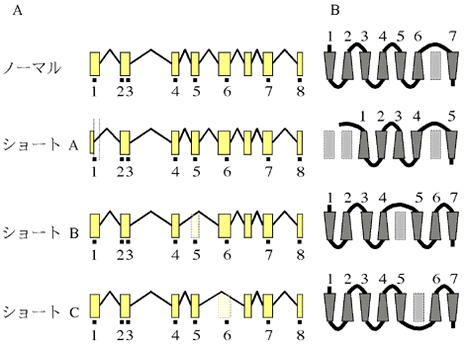 |
図3:選択的スプライシングと推定された膜貫通領域
A: 選択的スプライシングによる構成エクソンの変化。B: 推定された膜貫通領域の位置と方向を台形で表している。台形の上の数字は、TMの順番を表す。四角は、TM-PRでTMとしては使われていない位置を示す。
2. カルシウムイメージング法によるリガンドの特定
発現ベクター pIZT/V5(インビトロジェン)にナミアゲハのGPCR遺伝子PXFT-01989の配列を組み込み、エコーリン遺伝子を組み込んだベクターと同時に、鱗翅目昆虫であるヨトウガの卵巣組織由来のSf9細胞に導入した。このベクターはクローニングサイトとは独立にZeocin耐性遺伝子とGFP遺伝子が組み込まれているので、薬剤耐性とGFPの発現によって遺伝子が導入されていることを確認した。遺伝子導入細胞にセレンテラジンを浸透させるとエコーリンと結合し、GPCRの応答によってセカンドメッセンジャーとしてカルシウムが放出された場合に発光する。この手法により、細胞に導入したGPCRがどの化合物を受容するのかを確認することが可能になる。ミカンの葉から抽出した水溶性画分の混合化合物に対し、ノーマルタイプの遺伝子を導入した細胞とショートAタイプの遺伝子を導入した細胞で発光が確認されたので、10種類ある産卵刺激物質のうち購入または合成が可能な9種類について化合物単独での応答を観察した。その結果、どちらの導入細胞もシネフリンでのみ抽出混合化合物の場合と同様の反応が確認され、選択的スプライシングによって構造に変化が起きても機能は変わらない可能性が示唆された(図4)。今後、他のショートタイプ遺伝子を組み込んだ細胞の反応や、シネフリンに類似する化合物への応答など、より詳細な解析を必要するが、PXFT-01989のリガンドとしてシネフリンが有力な候補であると考えられる。
 |
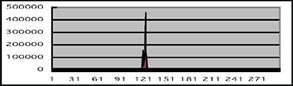 |
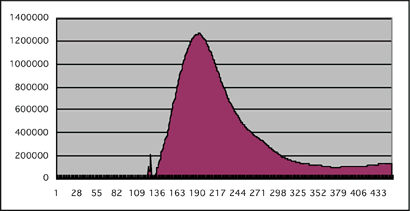
図4:導入細胞のカルシウム濃度変化
9種類の産卵刺激物質のうち、シネフリン投与でのみピーク型の反応が観察された。対象としてナリルチンに対する反応を示す。シネフリンでの最初の小スパイクと、ナリルチンでのスパイク状の反応は、化合物投与時の人為的な刺激によると考えられる。
3. シロオビアゲハからGPCRの部分配列を発見
ナミアゲハのGPCR遺伝子の配列をもとにプライマーを作成し、他のアゲハチョウのゲノムDNAをテンプレートとしてPCRを行った結果、シロオビアゲハからアミノ酸配列で65.6%の類似性を示す増幅産物が見つかった(図5)。この部分に特異的なプライマーを作成し、シロオビアゲハ雌成虫のふ節から調整したトータルRNAを用いRT-PCRを行った結果、増幅産物が確認されたため、この遺伝子はふ節で発現していることが明らかになった。RACE法で全長配列の決定を試みた結果、3’側についてはコード領域の全長配列が得られたものの、5’側については特定の部位で逆転写反応が終了し、全長配列を得ることができなかった。塩基配列を決定できた領域のゲノム配列を、ナミアゲハのGPCR遺伝子ゲノム配列とアライメントすると、イントロンが挿入されている部分が共通していた。また、どのイントロンもシロオビアゲハのものがナミアゲハのものより約2倍長くなっており、イントロンの部分には塩基配列の類似性はなかった。現在塩基配列が得られている範囲で、エキソン配列の相同性が高く、しかもエキソンがイントロンによって分断されている全ての部位が一致することは、両者が共通祖先から最近分岐したオルソログ関係にあることを示唆している。
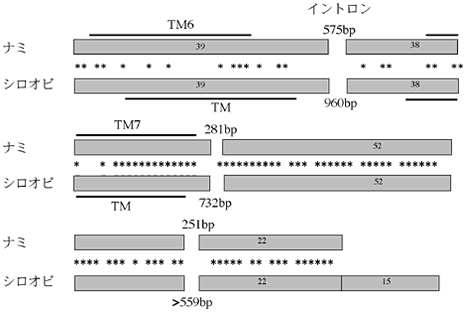 |
図5:CR:ス
アミノ酸配列の部分を矩形で模式した。矩形の中の数値は、その領域のアミノ酸数を表す。矩形の間の星印はアミノ酸が一致する部分を示す。TMは推定された膜貫通領域を表す。イントロンが挿入している位置は両者で一致する。C末端はシロオビアゲハのものが15アミノ酸長かった。イントロンは全て、シロオビアゲハの方が長かった。アライメントした151アミノ酸のうち、99アミノ酸 (65.6%)が一致した。
おわりに
ナミアゲハから見つかった産卵刺激物質レセプターと思われる遺伝子から、選択的スプライシングによって4種類のメッセンジャーRNAが作られており、1個の遺伝子から複数のレセプタータンパクが発現している可能性が示唆された。しかし、カルシウムイメージング法による機能解析の結果では、ノーマルタイプとショートAタイプが同じ化合物に反応していたことから、選択的スプライシングによる発現産物の多型があっても、機能には影響が無い可能性も考えられる。この遺伝子の機能の解明は、今後の我々の研究において最も重要な課題なので、詳細な解析を行いリガンドを特定する予定である。また、ショウジョウバエの感覚器官でこのGPCRを発現させて、ナミアゲハの産卵刺激物質への応答を観察するための準備、及び、ナミアゲハでRNAiにより発現を阻害した場合の電気生理的応答を観察するための準備を平行して進めている。これらの結果が得られれば、化合物を受容しシグナル伝達を引き起こすというレセプターとして機能していることの直接の証拠になると期待される。
シロオビアゲハからはナミアゲハ産卵刺激GPCR遺伝子と相同遺伝子の部分的な配列しか得られていないが、雌成虫の前脚ふ節で発現していることから、産卵刺激物質のレセプターとして期待される。ナミアゲハのGPCR遺伝子PXFT-01989とアミノ酸配列が類似するだけでなく、エキソンがイントロンによって分断されている位置など、遺伝子構造まで類似していることから、2種アゲハ間のオルソログ遺伝子である可能性が考えられる。この遺伝子の発見により、近縁種であるナミアゲハとシロオビアゲハの比較ゲノム的な研究を行うことへの道が開けた。さらに多くの産卵刺激GPCRを発見するために、新規に作成したcDNAライブラリーとゲノムライブラリーは、有力な道具になると思われる。
食草選択機構の変化は、アゲハチョウの仲間においては進化の原動力の一つであると考えられる。寄主転換が起きた際にはGPCRに何らかの変化があったことが予想されることから、このグループの進化に直接影響した可能性のある遺伝子の解明は、生物の進化という壮大な現象を解き明かす手がかりになることが期待される。来年度は、これらGPCR遺伝子の情報を利用して、他のアゲハからもこの遺伝子の探索を試みる予定である。
参考文献
1 Nishida R, Ohsugi T, Kokubo S, and Fukami H. (1987) Oviposition stimulants of a Citrus-feeding swallowtail butterfly, Papilio xuthus L. Experientia 43:342-344.
2 Feeny, P. (1995) Ecological opportunism and chemical constraints on the host associations of swallowtail butterflies, pp. 9-15. in J. M. Scriber, Y. Tsubaki, and R. C. Lederhouse (eds.). Swallowtail Butterflies: Their Ecology and Evolutionary Biology. Scientific Publishers, Gainesville, Florida.
3 Clyne PJ, Warr CG, Carlson JR. (2000) Candidate taste receptors in Drosophila. Science. 287:1830-1834.
4 Hill CA, Fox AN, Pitts RJ, Kent LB, Tan PL, Chrystal MA, Cravchik A, Collins FH, Robertson HM, Zwiebel LJ. (2002) G protein-coupled receptors in Anopheles gambiae. Science. 298:176-178.
5 Krieger J, Raming K, Dewer YM, Bette S, Conzelmann S, Breer H. (2002) A divergent gene family encoding candidate olfactory receptors of the moth Heliothis virescens. Eur J Neurosci. 16:619-628.
6 Galindo K, Smith DP. (2001) A large family of divergent Drosophila odorant-binding proteins expressed in gustatory and olfactory sensilla. Genetics. 159:1059-1072.
7 Ono H, Yoshikawa H. (2004) Identification of amine receptors from a swallowtail butterfly, Papilio xuthus L.; cloning and mRNA localization in foreleg chemosensory organ of host plants. Insect Biochem. Mol. Biol., 34: 1247-1256.