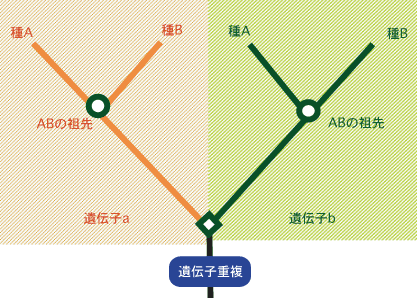
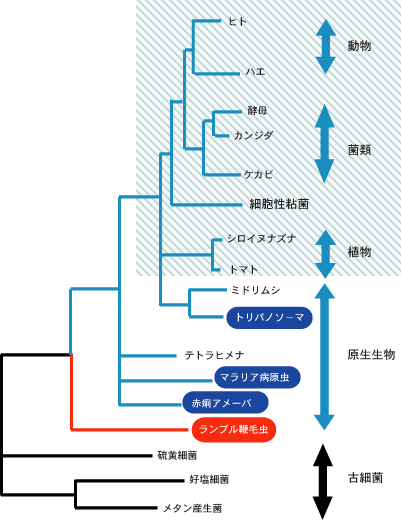
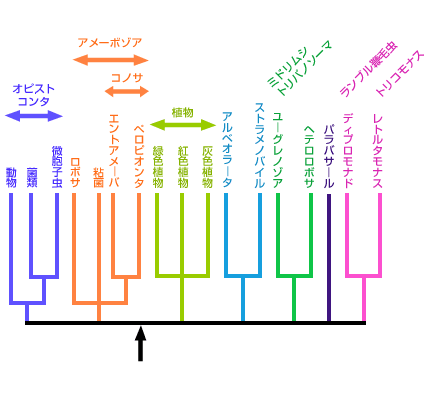
最新の研究やそれに関わる人々の話を交えて、生きものの進化に迫ります。 バックナンバー 【見直される真核生物の系統樹】 2006年1月16日 ミトコンドリアを持たない単細胞の原生生物、ランブル鞭毛虫とその他の真核生物との分岐が、真核生物の系統で起きた最初の大きな枝分かれであり、動物、菌類、植物のトリオ、すなわち、クラウングループは真核生物初期進化の最終段階で出現したというストーリーが、1990年代前半に確立し、以来ずっと信じられてきた。リボソームRNAやポリペプチド伸長因子などの単独の遺伝子からの系統樹推定に代わって、最近、多数の遺伝子に基づく解析が行われ、真核生物の系統進化が大きく見直され始めている。原生生物界動物界、植物界、菌界の関係多数の分子に基づく系統樹の推定紛らわしい原生生物:細胞性粘菌寄生性原生生物の系統的位置真核生物のルートはどこか? 原生生物界 カール・ウースの分類体系によると、地球上の全生物は、真正細菌、古細菌、真核生物の3つの超生物界に分類されるが、エドアール・シャトンの分類では、全生物は原核生物(ウースの分類では真正細菌と古細菌に対応する)と真核生物に分類される。もう一つ有名なのが1959年にロバート・ホイッタカーによって提唱された5界説である。5界説では、全生物は、モネラ界(原核生物に対応)、原生生物界(原生生物、藻類などの単細胞生物)、菌界(きのこ、かび、地衣植物など)、植物界(コケ類、シダ類、被子および裸子植物など)、および動物界(脊椎動物およびさまざまな無脊椎動物を含む多細胞動物)の5つの界に分類される。ホイッタカーに従って、真核生物超生物界は原生生物界、菌界、植物界、動物界の4つの界に分類される場合が多い。 原生生物は、大雑把には、単細胞の真核生物と考えて差し支えないのだが、なかには生活環に多細胞状態を含むものもあり、厳密な定義が難しい。リン・マーギュリスとカーリーン・シュヴァルツの名著、「五つの王国」には、原生生物とは、動物でもなく、植物でもない、さりとて菌類でもない、しかも原核生物でない生物とある。こう言ってしまうとわけの分からない生物が一緒くたにかき集められた、いわばごみの捨て場の生物界といった感じになるが、実際は多様で厳密な定義が難しいということなのであろう。 原生生物の形態はさまざまで、正確な種の数は定かでないが、ジョージ・メリンフェルドによれば、6万5千種以上にものぼる。ジョン・コーリスの推定では25万種にもなると「五つの王国」に書いてある。この多様な原生生物界も幾つかの門に分類されている。例えば、「五つの王国」によると、30の門から構成されている。実のところ、原生生物については、分類に関しても、系統に関しても、いまだにいろいろと議論の多い生物界である。 原生生物の中で分子生物学的研究が進んでいるグループは、睡眠病やマラリア原虫など、病原性のある生物や寄生性の生物で、医学的興味から研究が進んでいる例外的な原生生物で、これほど多様な生物界にしては分子レベルの研究が進んでいるグループがあまりにも少ない。分子レベルの研究が遅れている理由として、マーギュリスとシュヴァルツは、原生生物のほとんどが食物にならず、直接経済的な重要性も持っていないので、この分野の研究に経済的な投資がされてこなかったからだと述べている。 ▲このページのトップへもどる 動物界、植物界、菌界の関係 花が好きなのは昆虫に限ったことではない。花は嫌いだという人はめったにいないであろう。逆に、風呂場の下水にこびりついた黒カビが好きだという人もお目にかかったことがない。われわれは花に近いのか、それともカビに近いのか?と聞いたら、心情的には花と言いたいところであろう。真核生物超生物界で原生生物界を除く3つの界は多細胞の形態を取るが、花は植物界に属し、カビは菌界で、われわれはいうまでもなく動物界に属している。従って上の質問は、学問的に言い直せば、3つの生物界の系統関係を問うていることになる。 この問題は身近で、誰もが興味を持つ問題で、しかも学問的にも重要な問題だったために、分子進化学がスタートして間もない1970年代には早くも分子系統学的研究がスタートしている。化石のデータが極めて少ないので、形態レベルでの研究が立ち後れていたこともあって、分子レベルでの研究が乗り出していく良い機会であったのであろう。最初にこの問題を手がけた分子進化学者はマーガレット・デイホッフであった。彼女はチトクロームcというミトコンドリアで働くタンパク質を使って、彼女自身が開発した系統樹推定法で解析し、動物と菌類は近縁で、植物は両者に対して遠縁の関係にあることを示した。この系統関係を動物-菌類近縁系統樹と呼んでおこう。ちなみに三者の可能な関係には、このほか、動物と植物が近縁で菌類が両者に対して遠縁の関係(動物-植物近縁系統樹)と、植物と菌類が近縁で、動物が遠縁の関係(植物-菌類近縁系統樹)がある。 翌年、ウオルター・フィッチは同じタンパク質を使って、しかし方法は自分自身で開発した方法によって、動物と植物が近縁で、菌類はそれらより遠縁の関係、動物-植物近縁系統樹、を推定した。我が国では長谷川政美(現、統計数理研究所教授)が早くから一貫して精度の高い最尤法でこの問題に取り組み、今では当たり前のようにして利用している最尤法を中心に、分子系統樹推定法の確立に大きな成果を残している。それ以来1995年までに、いろいろな分子で、いくつもの異なる方法で解析が行われ、20を下らない論文が報告されている。こうした論文では、著者ごとに異なる系統樹が主張されている。 なぜ研究者によって、これほどまでに違う答えが得られるのであろうか?理由は幾つか考えられる。3つの生物界の分岐は非常に古い時代に起きているため、分子によっては,アミノ酸や塩基が変化できる座位は目一杯変わってしまっている(変化が飽和しているという)ため、正しい進化の情報が得られないことが考えられる。あるいは、3つの生物界の分岐時期が比較的接近しているために、小さな分子ではアミノ酸置換数の十分な統計量が得られず、精度よく3つの樹形が分離できなかったことも考えられる。使用する分子や方法によっていくぶん結果が違ってくることは十分予想されることだが、三者の間で起きた分岐の時期があまりにも接近しているので、分子や方法の違いによるバラツキの範囲内に収まってしまっている可能性がある。 ▲このページのトップへもどる 多数の分子に基づく系統樹の推定 こうした困難を克服するために、二河成男(現、放送大学助教授)を中心とする著者のグループは、これまで1種類の遺伝子で推定されていた動物・植物・菌類の系統関係に対して、23種類の異なるタンパク質を用意し、また系統樹の推定法についても、異なる3つの方法を併用して解析を行った。さらに一つ一つのタンパク質で得られた結果を重ね合わせ、総合的に判断できるような統計処理を施した。一つ一つの分子は小さいため、結果がばらつくが、全部重ねると大きな分子で解析したことと同じことになるので、誤差が小さくなると期待できる。 ここで、たくさんの遺伝子(あるいは分子)を使って生物の系統樹を推定する際に注意しなければならない点を指摘しておこう。一般に生物の種の分岐に際し、それぞれの種に同じ遺伝子が受け渡される。別の言い方をすれば、種の分岐と同時に遺伝子も分岐することになる。この場合、種の分岐と遺伝子の分岐は対応すると期待される。分子から生物の系統を推定するときに用いる遺伝子はこのタイプの遺伝子でなければならない。このように、種の分岐と遺伝子の分岐が対応している関係を「オーソログ」という。 いま、共通の祖先から2つの種A、Bが分岐したとしよう。この種分岐に先行して遺伝子重複が起きている場合を考えてみよう(図1)。遺伝子重複で生じた2つの遺伝子をa、bとしよう。遺伝子重複が種分岐に先行して起きているので、それぞれの種は2つの重複遺伝子a、bを持つ。このとき種Aの遺伝子aと種Bの遺伝子a、あるいは種Aの遺伝子bと種Bの遺伝子bはオーソロガスの関係にあることになり、種の分岐と遺伝子の分岐が対応している。一方、種Aの遺伝子aと種Bの遺伝子b、あるいは種Aの遺伝子bと種Bの遺伝子aでは、いずれも種の分岐と遺伝子の分岐が対応していない。このような関係のことを「パラログ」の関係という。パラログな関係にある遺伝子あるいはタンパク質を使って系統樹を推定すると、正しい系統関係が得られない。動物・植物・菌類に対して、いずれもオーソログの関係にある分子を使った場合と、どれか一つにパラログの関係にある分子がまぎれこんだ場合とでは、推定された系統樹が一般に違ってくる。 図1. 遺伝子重複とパラログ、オーソログの関係 23種類のタンパク質のデータから作った組には、パラログのデータを持つ組があるかも知れないが、その数が非常に少なければ、大勢にはあまり影響することがないであろう。パラログのデータを持つ組は、他の組と比べて、著しく違った結果を示すので、そうした組を除外することもできる。 結果は明瞭であった。動物は菌類に近縁で、植物に遠縁の関係にあることがはっきりと示された。3つの方法で同じ結果を示すタンパク質に限定すると、いっそう明瞭な結果が得られた。また、パラログのデータを含んでいると思われる組を除外した場合でも、同じ結果が得られた。こうして3つの生物界の系統関係に関する混乱が収拾された。この研究からわれわれが学んだ重要な点は、分子から生物の間の系統関係を明らかにしようとしたら、できるだけたくさんの分子を使うべきで、一つの分子から得られた結果から結論を出すのはあまりにも危険すぎるということだ。 ▲このページのトップへもどる 紛らわしい原生生物:細胞性粘菌 こうして動物・植物・菌類の系統関係は一段落したが、ここに細胞性粘菌という名の紛らわしい原生生物がいる。この生物の分類をめぐって、生物学者の間で論争が絶えない。この粘菌は、動き回り、ある時は集まってナメクジのようにもなり、変態までする。またバクテリアを食べて消化することもできる。まるで動物のようである。一方で、この粘菌は植物のように、まっすぐな体になって、実をつけ、胞子を作る。そうかと思えば、湿度の高い土壌や腐敗した植物から栄養を摂取する。また胞子は頑丈な細胞壁を持っている。これらの点では菌類によく似ている。動物学者によっては、動菌類と呼ぶ人もいる。動物界にも、植物界にも、菌界にも入れられないので、がらくた生物界である原生生物界に放り込まれたわけである。 形態レベルで見るとそんなに分類が紛らわしいのなら、分子で調べてみる価値がある。隈啓一(現、京都大学化学研究所助教授)が中心になって、著者のグループで解析が始まった。当時用意できたタンパク質の配列データ数は19種類と十分な数であった。このタンパク質データセットで、動物界、植物界、菌界に加え、粘菌の間の系統樹が決定された。その結果は図2に示したように、細胞性粘菌は動物と菌類の共通の祖先と枝分かれし、植物とは遠い関係にあることがはっきりした。最近、細胞性粘菌の全ゲノム塩基配列が決定し、多くの遺伝子でこの系統関係が再検討された。その結果、上記の結果が再現された。 図2. 1990年代に支持されていた真核生物の系統樹. ポリペプチド伸長因子で推定された系統樹. クラウングループを含む系統を網掛けで、寄生性の原生生物を反転で示した。 植物界の起源に関しては一般に意見が一致している。すなわち、緑色陸上植物は緑藻から生じたと考えられている。分子のデータも同じ結論に達している。菌界の起源については、まだはっきりしていない。動物の起源に関しては、最近研究が進んでおり、かなりはっきりしてきた。 ▲このページのトップへもどる 寄生性原生生物の系統的位置 分子から生物の系統を明らかにしようとする場合、まず配列データが得やすいリボソームRNAで解析を始めるのが通常のパターンである。次いでタンパク質を使った解析へと進めるのだが、この場合も配列データが簡単に得られるポリペプチド伸長因子を使う場合が多い。真核生物の初期進化の研究も例外ではなく、リボソームRNAでスタートし、ついでポリペプチド伸長因子で前者の結果を確認するという手順で研究が進められてきた。1990年代前半に解析され、今日まで広く信じられてきた系統樹が、若干の食い違いがあるものの、図2のような系統樹である。 動物・菌類・植物の系統関係に関しては、1980年代後半までに研究がかなり進み、上で述べたように、多数の遺伝子を使って生物の系統を再現する研究が1990年代前半までにすでに行われていたわけである。ただし、この場合、解析に含めた生物種はこれら3つのグループに限定されていた。他の原生生物は3つのグループの外群になるか、あるいはそれぞれへ至る枝の途中から枝分かれすると期待されていた。実際どのような分岐パターンを示すかは、多数の遺伝子を用いて、多くの主要な真核生物のグループを含む系統解析が待たれていた。 ところで、図2の系統樹の大きな特徴は、寄生性のグループが真核生物の系統樹上非常に古い時期に分岐している点である。これは、系統樹推定にしばしば見られる”Long Branch Attraction Artifacts“: LBAと呼ばれている人為的結果ではないかと指摘されていた。分子系統樹では、ある生物が他の生物と分岐後現在に至る間に蓄積した塩基あるいはアミノ酸置換の数に比例して系統樹の枝の長さを表現する慣わしになっている。非常に古い時期に分岐した生物は、分岐後経過した時間が長いので、系統樹では枝の長さが最近分岐した生物に比べで長い。しかし、比較的最近分岐した生物でも、進化の速度が非常に高い場合には置換数が多くなり、誤って古い時期に分岐したように推定されてしまう場合がある。さらに、研究の初期にしばしば利用されたリボソームRNAやポリペプチド伸長因子は、進化速度がときどき系統間で大きく変動する場合があることが知られている。こうしたことがLBAを助長した可能性が大いにある。 微胞子虫の系統的位置は、リボソームRNAやポリペプチド伸長因子でLBAの結果、誤って非常に古い時期に推定されていた典型的な例である。微胞子虫類は「五つの王国」によると、極_胞子虫門の微胞子虫綱(ミクロスポラ)に分類される寄生性原生生物である。微胞子虫の仲間にはグルゲアやエンセファリトズーンなどがあり、宿主細胞内で病原体として生きているものが多い。これら微胞子虫の仲間は、典型的なミトコンドリアやペルオキシソームといった細胞小器官をもたない。加えて、リボソームRNAやポリペプチド伸長因子による分子系統樹から、微胞子虫は真核生物の中で最も古い時期に分岐したグループであることが示されたことから、一時、微胞子虫はミトコンドリアの共生が起こる以前の真核生物、すなわち、アーケゾアではないかと騒がれた(本シリーズ;真核生物誕生の謎を参照)。 しかし、微胞子虫はミトコンドリアで働く熱ショックタンパク質HSP70が核のDNAに存在することが明らかになったことから、かつてミトコンドリアを持っていたが、おそらく寄生性の獲得の結果、ミトコンドリアを二次的に失ったのではないかと考えられるようになった。ミトコンドリアが消失する以前にミトコンドリアDNA上の遺伝子の一部を核DNAに移したというわけである。最近、HSP70の抗体を作ることで、微胞子虫がミトコンドリアを完全に消失したのではなく、残存構造が存在することが示された。さらに、RNAポリメラーゼIIやいくつかの分子に基づく系統解析は、微胞子虫が驚くことに菌類に近縁であることを明らかにした。こうして、動物、菌類、微胞子虫は系統的に、一つのまとまった大きなグループを形成することが明らかになり、オピストコンタと名づけられた。 さらに多数の遺伝子を用いた系統解析から、多様なアメーバのグループ、アカントアメーバ(ロボサに分類)、コノサに分類されるエントアメーバ、マスティグアメーバ(ペロビオンタに分類)、及び細胞性粘菌が、一つの大きなグループ、アメーボゾアに分類された。最近では、上記のオピストコンタ、アメーボゾアに加えて、7つから8つの大きな真核生物のグループが確認されている(図3)。すなわち、繊毛虫(ゾウリムシ、テトラヒメナなど)、渦鞭毛藻類、アピコンプレックス(マラリア原虫など)から成るアルベオラータは、珪藻類、褐藻類、卵菌類などのストラメノパイルと近縁な関係にあって、一つの大きなグループを作っている。ミドリムシやトリパノソーマを含むユーグレノゾアは、ナエグレリアやアクラシスなどのヘテロロボサと姉妹群である。さらに、ディプロモナド(ランブル鞭毛虫など)とレトルタモナス(トリコモナスなど)は近縁関係にある。 ▲このページのトップへもどる 真核生物のルートはどこか? さて、ここまで話を進めてきたが、最後に重大な問題を考えてみよう。動物、植物、菌類をまとめてクラウングループと呼んでいるが、これらの系統は本当に一つにまとまった単系統群を形成するのであろうか?これらの系統は全真核生物の系統樹のどこに位置するのであろうか?全真核生物の系統樹の最も深い根(ルート)はなにか?同じ問だが、真核生物のなかで最古の分岐はなにか?こうした問に答える研究は1990年代に入って盛んに行われてきたが、最近、見直しが行われ始めている。ひょっとすると従来の系統樹が根底から覆る可能性が出てきているのだ。 橋本哲男(筑波大学教授)らは、上で述べた真核生物の大きな分類群の間の系統関係を明らかにする目的で、多数の分子を用いて分子系統解析を試みた。しかし、残念ながら統計的に有意な結果は得られなかった。一方で、ステックマンとカバリエ・スミスは異なる視点、すなわち、分子的な共有形質でこの問題の解明を試みている:2つの鞭毛を持つ植物やアルベオラータなどのグループ(バイコント)では、デハイドロフォレートレダクターゼ(DHFR)とチミジレートシンターゼ(TS)がゲノムの上で逆向きに融合している。一方、オピストコンタとアメーボゾアでは2つの遺伝子は融合していない。ちなみに2つのグループは一つの鞭毛を持ち、ユニコンタと名づけられている。彼らはさらに、ピリミジン合成系に含まれる3つの遺伝子の遺伝子融合についても調べた。その結果、ユニコンタでは遺伝子融合が見られたが、バイコンタでは見られなかった。これらの結果からかれらは、真核生物の最も深い根は植物へ至るバイコンタのグループとオピストコンタとアメーボゾアを含むユニコンタのグループの間にあると示唆した。 彼らの結果は真核生物の最も深いルートについての重要な示唆を与えてはいるが、こうした分子的形質の保存性に関する十分なデータの蓄積がないので、信頼性が十分あるとは断言できない。最終結論までにはより多くのデータの蓄積が重要であろう。また、最近では遺伝子内の特定のイントロンの有無で同様の解析を行う試みがあるが、この種類の解析は注意深く行うべきである。おそらく寄生性の生物では高速でのゲノム複製の結果、ゲノムサイズが極端に小さくなり、イントロンが選択的に欠落している可能性が十分ある。これはまた、LBAの結果と類似の傾向を示す。 現時点では不十分な証拠ではあるが、全真核生物は、大きくオピストコンタとアメーボゾアからなる大グループ(動物大グループと呼ぶことにする)と、図3でオピストコンタとアメーボゾア以外のグループからなる大グループ(植物大グループ)に分けられ、最も深い根が両大グループの間に位置する可能性があるかも知れない(図3の矢印の位置)。もし、この系統樹が正しいなら、1)動物・菌類・植物からなるクラウングループは、これまで言われてきたような、真核生物初期進化の最終段階で出現した単系統群ではなく、真核生物の進化の最初期に分岐したことになる。2)このことはさらに、動物へ至る系統と植物へ至る系統で独立に多細胞化が起きたことを意味する。3)多細胞生物は、真核生物の進化の比較的早い時期に出現した可能性まである。こうした興味深い問題を含んだ真核生物の系統樹が、おもに多数の遺伝子を使った系統解析によって現在見直されつつある。 図3. 分子で推定された真核生物の大きな分類群. 矢印は一つの可能なルート. ▲このページのトップへもどる [宮田 隆] 宮田 隆の進化の話 最新号へ